КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ

![]()
|
КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
Глава VI. Систематика Hexapoda: общая ХАРАКТЕРИСТИКА и первичнобескрылые
VI-1. Hexapoda
Latreille 1802 – шестиножки, или насекомые
(Somobialia
Eucaryota
Metazoa
Monostomata
Haemataria
Peritoneata Metameria
Gnathopoda
Euarthropoda
Mandibulata
Atelocerata Hexapoda)
(рис. 6.1.1–10.16.2)
|
Название, однозначно соответствующее по циркумскрипции: — Hexapoda Latreille 1802a (T.2: 344-345) (non Hexapoda Batsch 1789; nec Hexapoda Latreille 1796). NomCirc. По циркумскрипции также соответствует: — «Insecta» sensu Leach 1815a (non Linnaeus 1758). Иерархическое название: Pediculus/f3=Scarabaeus/g3 [f:1777; g:L.1758(gen.170)] (sine Scolopendra; incl. Podura).
|
|
Ранг. От подкласса до подтипа, обычно класс или надкласс.
Аутапоморфии Hexapoda
● Постоянное число сегментов и эпиморфоз. Число посторальных сегментов постоянно и равно 17; из них первые 4 сегмента входят в состав головы, следующие 3 сегмента образуют грудь, и следующие 10 сегментов образуют брюшко (исключение составляют ногохвостки и бессяжковые, у которых число сегментов брюшка уменьшено – см. ниже, Ellipura); все 17 сегментов формируется при эмбриональном развитии под яйцевыми оболочками и их число остается неизменным в процессе постэмбриогенеза, т.е. имеет место эпиморфоз. Число обособленных ганглиев вентральной нервной цепочки [которое исходно соответствует числу сегментов – см. Metameria) у насекомых всегда меньше семнадцати: помимо того, что первый посторальный ганглий (тритоцеребрум) вошел в состав надглоточного ганглия (см. Euarthropoda), а II–IV посторальные ганглии, иннервирующие ротовые придатки головы, слиты в подглоточный синганглий, из десяти сегментов брюшка только первые 8 имеют собственные ганглии, тогда как IX и X сегменты брюшка иннервируются от ганглия VIII сегмента брюшка. Во многих таксонах насекомых произошло дальнейшее уменьшение числа ганглиев за счет слияний тех или иных ганглиев груди и/или брюшка.
Преоральная область и 17 посторальных сегментов, составляющих тело насекомого, дифференцированы следующим образом.
Голова (caput) у Hexapoda состоит из преорального отдела, несущего антенны, фасеточные глаза и глазки (см. Euarthropoda), и четырех посторальных сегментов (I–IV сегменты). Поскольку первый посторальный сегмент лишен конечностей (см. Atelocerata), голова несет 3 пары посторальных конечностей, служащих ротовыми придатками – мандибулы, максиллы и нижнюю губу. Поверхность головы утратила разделение на сегменты и представляет собой цельную головную капсулу – краниум (cranium). Исходное положение головы гипогнатное, т.е. направленное ротовыми придатками вентрально, так что мандибулы расположены впереди максилл, а максиллы – впереди нижней губы, в соответствии с исходным расположением их сегментов; в ходе эволюции положение головы многократно менялось на прогнатное (т.е. ротовыми придатками вперед, так что мандибулы расположены дорсально, а нижняя губа вентрально) и обратно на гипогнатное. Ротовой аппарат Hexapoda состоит из непарной верхней губы (см. Euarthropoda), пары одночлениковых мандибул (см. Mandibulata и Atelocerata), пары максилл сложного строения (см. ниже), пары суперлингв (см. Mandibulata), гипофаринкса (прасположенного между максиллами – см. Mandibulata) и нижней губы сложного строения (см. ниже). Исходно ротовой аппарат грызущий: мандибулы служат для разгрызания пищи, максиллы манипулируют частицами пищи и перемещают их в рот, нижняя губа удерживает пищу снизу, а телоподиты максилл и нижней губы служат осязательными органами – щупиками. Исходной пищей насекомых являются детрит, грибы, водоросли, споры и пыльца растений. В некоторых таксонах Hexapoda ротовой аппарат преобразовался из грызущего в иной – сосущий, колюще-сосущий, лижуще-сосущий, фильтрующий, скребущий и др., причем в каждом случае произошли уникальные преобразования или утрата определенных частей ротового аппарата. Некоторые насекомые приобрели ту или иную пищевую специализацию – хищничество, фитофагию или иную. В большинстве случаев нет однозначной связи между модификацией ротового аппарата и пищевой специализацией.
Грудь, или торакс (thorax), Hexapoda состоит из трех сегментов (V–VII посторальные сегменты – см. ниже), несущих по паре ног особого строения (см. ниже).
Брюшко, или абдомен (abdomen) у различных Hexapoda имеет настолько разнообразное строение, что до недавнего времени не было ясного представления о числе входящих в него сегментов и об их исходном строении. По новым данным брюшко гексапод состоит из 10 сегментов, являющихся VIII–XVII посторальными сегментами тела. Первые 9 из этих сегментов имеют повторяющиеся черты строения (см. ниже), а десятый сегмент брюшка имеет особое строение. У многих насекомых число сегментов брюшка кажется меньшим десяти, так как последние сегменты бывают сильно модифицированы, уменьшены и/или втянуты в предыдущие; лишь у ногохвосток и протур число сегментов брюшка действительно меньше десяти (см. ниже, Ellipura).
Синонимом термина «абдомен» является термин «уросома» (urosoma); для краткости сегменты брюшка насекомых называют уромерами, а их тергиты и стерниты, соответственно, уротергитами и уростернитами.
● Максиллы. В ротовом аппарате, помимо мандибул (специализированных жующих конечностей II посторальной пары – см. Mandibulata) имеется еще одна пара подвижных специализированных жующих конечностей – нижних челюстей, или максилл (maxillae), т.е. конечности III посторальной пары, вовлеченные в ротовой аппарат (см. Mandibulata). Строение максилл Hexapoda является уникальным и едино как для Entognatha (рис. 6.3.1E), так и для Amyocerata (рис. 6.7.1D, 9.1.3F).
Строение максиллы (рис. 6.7.1D, 9.1.3F). Проксимальная часть максиллы, входящая в состав стенки головы, разделена на две подвижно сочлененные части – поперечно направленный кардо (cardo) и продольно направленный стипес (stipes); каждая из этих частей не является самостоятельным члеником и непосредственно причленена к голове своей длинной медиальной стороной; от заднего (вентрального) края каждой из них идут мышцы к эндоскелету головы – к фулькруму (см. ниже, Entognatha) или к тенториуму (см. ниже, Amyocerata); в случаях утраты тенториума эти мышцы крепятся непосредственно к головной капсуле.
Выступающими частями максиллы являются лациния, галеа и щупик. На дистальном конце стипес несет жевательную долю – лацинию (lacinia), от основания которой отходят две мышцы, идущие впереди (дорсальнее) мышц стипеса и кардо: одна из мышц лацинии идет в стипес, другая – к головной капсуле. Исходно лациния склеротизована, заострена и зазубрена. Латеральнее и дорсальнее лацинии от стипеса отходит другой придаток – галеа (galea); от ее основания идет мышца в стипес, эта мышца лежит позади (вентральнее) всех прочих мышц стипеса. Исходно галеа не склеротизована, туповершинная. Латеральнее и дорсальнее галеи от стипеса отходит максиллярный щупик (palpus maxillaris), от основания которого мышцы идут в стипес. Исходно щупик расчленен, и в каждом его членике кроме последнего имеется одна или несколько мышц, двигающих следующий членик [см. (2c)].
Благодаря тому, что проксимальная часть максиллы имеет два подвижно сочлененных склерита (кардо и стипес), к каждому из которых подходят мышцы, максилла способна совершать сложные движения, двигаясь вперед-назад и медиально-латерально; в результате этого лацинии максилл, вооруженные зубцами и щетинками, способны манипулировать частицами пищи и загребать их в рот.
У некоторых Hexapoda максиллы модифицированы так, что утратили те или иные части, иногда максиллы совсем утрачены.
У Entognatha и Triplura максиллы сохраняют кардо, стипес, лацинию, галею и щупик. У личинок поденок максиллы сохраняют кардо, стипес, лацинию и, как правило, щупик, но утратили обособленную галею (см. ниже, Ephemeroptera). У стрекоз максилла утратила либо галею, либо щупик (см. ниже, Odonata). У Idioprothoraca, Rhipineoptera и Zoraptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик. У Acercaria максиллы утратили кардо и подвижную галею, а стипес и лациния сильно преобразованы; щупик сохраняется или утрачен (см. ниже, Acercaria, Condylognatha, Thysanoptera, Arthroidignatha). У большинства жуков (Eleuterata) максиллы сохраняют кардо, стипес, лацинию, галею и щупик. У Strepsiptera максиллы вестигиальные, как и весь ротовой аппарат (см. ниже, Strepsiptera). У имаго всех Neuropteroidea и у личинок Rhaphidioptera и Meganeuroptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик; при этом у личинок вислокрылок максиллы по-разному модифицированы (см. ниже, Nothomegaloptera и Eumegaloptera). У личинок Birostrata максиллы, вошедшие в состав парного колюще-сосущего ротового аппарата, уникальным образом модифицированы (см. ниже, Birostrata). У Hymenoptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик, бывают по-разному модифицированы у личинок и имаго. Среди Enteracantha, у личинок и у имаго Scorpiomuscae, Nannomecoptera, Metamecoptera и Raphioptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик; у имаго блох максиллы сильно модифицированы, без кардо и галеи, с погруженным стипесом, колющими лациниями и модифицированным щупиком (см. ниже, Aphaniptera). У личинок Diptera максиллы либо сохраняют кардо, стипес, лацинию, галею и щупик, либо имеют упрощенное строение; у имаго двукрылых галеа утрачена, а максиллы исходно специализированы как колюще-режущие (см. ниже, Diptera); у имаго Cyclorapha и некоторых других максиллы сохранили только щупики, тогда как их остальные части вошли в состав непарного хоботка (см. ниже, Cyclorapha). Среди Amphiesmenoptera, у личинок и у имаго Trichoptera и Protolepidoptera максиллы сохраняют кардо, стипес, лацинию, галею и щупик; у имаго Glossolepidoptera максиллы утратили лацинии, а их галеи преобразовались в сосущий хоботок (см. ниже, Glossolepidoptera).
Число члеников максиллярного щупика. По-видимому, исходным для Hexapoda является наличие 5 члеников в максиллярном щупике (в отличие от 3-членикового лабиального щупика – см. ниже). Иногда число члеников уменьшено благодаря редукции или слиянию тех или иных члеников, иногда щупики утрачены; реже число члеников щупика увеличено благодаря вторичному расчленению некоторых члеников.
У Zygentoma максиллярные щупики 5-члениковые. У Microcoryphia максиллярные щупики сильно удлинены и число их члеников увеличено до 7 (см. ниже, Microcoryphia). У поденок и стрекоз число члеников максиллярного щупика различным образом сокращено (см. ниже, Ephemeroptera и Odonata). У всех Idioprothoraca и большинства Rhipineoptera максиллярные щупики 5-члениковые (в единичных случаях 4-члениковые). Среди Parametabola 5-члениковые максиллярные щупики сохраняются у Zoraptera. У Acercaria максиллярные щупики имеют сокращенное число члеников: имеется не более 4 члеников, лишь у некоторых трипсов щупики вторично расчленены.
У многих Metabola по крайней мере на стадии имаго максиллярные щупики 5-члениковые; у личинок Metabola максиллярные щупики обычно также 5-члениковые, но значительно меньшего размера, с сильно укороченными члениками. У имаго и личинок жуков число члеников максиллярного щупика, вероятно, исходно уменьшено до 4, иногда сокращено до 3 или 2, реже вторично увеличено до 5. У имаго и личинок веерокрылых максиллы сильно видоизменены (см. ниже, Strepsiptera). У имаго сетчатокрылообразных максиллярные щупики обычно 5-члениковые, лишь у некоторых Eumegaloptera 4-члениковые; у личинок Birostrata максиллярный щупик не расчленен и уникальным образом модифицирован (см. ниже, Birostrata); у личинок других сетчатокрылообразных (Rhaphidioptera и Meganeuroptera) максиллярный щупик имеет то же число члеников, что у имаго. У имаго перепончатокрылых число члеников максиллярного щупика, вероятно, исходно увеличено до 6; бывает от 1 до 6 члеников; у личинок перепончатокрылых максиллярный щупик 5-члениковый или с меньшим числом члеников. У имаго всех Enteracantha (скорпионницы и блохи) максиллярные щупики 5-члениковые (у блох кажутся 4-члениковыми – см. ниже, Aphaniptera); у личинок число члеников максиллярного щупика уменьшено до 3 (у большинства скорпионниц) или до 2 (у Calyptroptera). У имаго Diptera максиллярные щупики исходно 5-члениковые, в некоторых таксонах число члеников сокращено; в частности, у Brachocera число члеников сокращено до двух, у Brachocera-Cyclorapha – до одного (см. ниже, Brachocera и Cyclorapha); у личинок двукрылых максиллярные щупики обычно нерасчлененные. У имаго Trichoptera максиллярные щупики исходно 5-члениковые; у некоторых представителей модифицированы, с уменьшенным числом члеников; иногда вестигиальные или утрачены; у личинок Trichoptera максиллярные щупики 4-члениковые. У имаго Lepidoptera максиллярные щупики исходно 5-члениковые, но у большинства представителей вестигиальные, с уменьшенным числом члеников, или утрачены (см. ниже, Lepidoptera); у личинок Lepidoptera максиллярные щупики имеют не более 3 члеников.
● Нижняя губа. Конечности IV посторальной пары (соответствующие максиллам II многих Eucrustacea – см. Mandibulata) слились медиально своими коксоподитами, образовав непарную нижнюю губу, или лабиум (labium) (рис. 7.2.3H); нижняя губа является третьей парой ротовых конечностей и последней парой конечностей головы. Гипофаринкс (непарный выступ между основаниями максилл – см. Mandibulata) оказался расположенным между ротовым отверстием и нижней губой. Протоки, идущие от пары желез, принадлежащих IV посторальному сегменту, обычно соединяются в непарный проток, который открывается непарным отверстием у основания нижней губы, обычно между нижней губой и гипофаринксом; эти железы называют лабиальными (glandulae labiales), или слюнными (g. salivares).
Строение нижней губы. Нижняя губа дистально несет две пары отчлененных придатков: более медиальные придатки – глоссы (glossae) и более латеральные придатки – параглоссы (paraglossae); латеральнее параглосс причленяется пара членистых лабиальных (нижнегубных) щупиков (palpi labiales). К основанию каждой глоссы, параглоссы и щупика подходит одна или несколько мышц, неподвижные концы которых крепятся в непарной части нижней губы. Внутри глосс и параглосс мышц нет. Щупик обычно расчленен, и в каждом его членике кроме последнего имеется одна или несколько мышц, двигающих следующий членик. У некоторых Hexapoda глоссы и/или параглоссы слиты или редуцированы.
У Entognatha нижняя губа неподвижная, так что глоссы и параглоссы слиты, щупики одночлениковые или утрачены (см. Entognatha). У Triplura глоссы и параглоссы развиты, причем у Microcoryphia параглоссы расщеплены на доли (см. ниже, Microcoryphia); щупики развиты. У личинок Ephemeroptera глоссы, параглоссы и щупики обычно развиты, но в некоторых таксонах глоссы и/или параглоссы слиты, а их мышцы утрачены (у имаго Ephemeroptera ротовой аппарат утрачен). У стрекоз нижняя губа сильно модифицирована, глоссы и параглоссы утрачены, а щупики модифицированы (см. ниже, Odonata). У Idioprothoraca (эмбии и гриллоблаттиды) глоссы, параглоссы и щупики развиты. У большинства Rhipineoptera глоссы, параглоссы и щупики развиты; у уховерток глоссы утрачены и имеются лишь параглоссы (см. ниже, Dermatoptera). У Zoraptera нижняя губа дистально разделена на пару долей, каждая из которых подразделена на две доли, вероятно соответствующие глоссе и параглоссе; щупики развиты. У всех Acercaria глоссы и параглоссы плохо выражены или отсутствуют; при этом у Parapsocida, Eupsocida, Thysanoptera и части Mallophaga щупики имеются, у прочих Parasita и у Arthroidignatha все парные придатки нижней губы утрачены (см. ниже, Acercaria). У имаго и личинок насекомых с полным превращением по крайней мере парные глоссы отсутствуют: они либо слиты вместе, либо совсем утрачены; парные параглоссы бывают развиты или отсутствуют; щупики обычно развиты.
У большинства Hexapoda непарная часть нижней губы расчленена на проксимальную часть – постментум и дистальную часть – прементум. Постментум (postmentum) [называемый также субментум (submentum)] обычно имеет только заднюю (вентральную) стенку. Прементум (praementum) [называемый также ментум (mentum)] представляет собой непарный придаток, подвижно причлененный к переднему краю постментума; глоссы, параглоссы и щупики причленены к прементуму, а идущие от их оснований мышцы крепятся внутри прементума.
Число члеников лабиального щупика. По-видимому, исходным для Hexapoda является наличие 3 члеников в лабиальном щупике (в отличие от 5-членикового максиллярного щупика). Иногда число члеников лабиального щупика уменьшено благодаря редукции или слиянию тех или иных члеников, иногда щупики утрачены; реже число члеников щупика увеличено благодаря вторичному расчленению некоторых члеников.
У Entognatha лабиальные щупики 1-члениковые или утрачены (см. ниже, Entognatha). У Zygentoma лабиальные щупики выглядят 4-члениковыми (см. ниже, Zygentoma). У Microcoryphia лабиальные щупики 3-члениковые. У личинок поденок лабиальные щупики обычно 3-члениковые, иногда 2-члениковые. У стрекоз нижняя губа сильно модифицирована, и ее щупики утратили расчленение (см. ниже, Odonata). У Idioprothoraca и Rhipineoptera лабиальные щупики 3-члениковые. Среди Parametabola 3-члениковые лабиальные щупики сохраняются у Zoraptera. У Acercaria лабиальные щупики подверглись редукции: они имеют не более 2 члеников, и лишь у некоторых трипсов вторично расчленены.
У многих Metabola трехчлениковые лабиальные щупики имеются по крайней мере на стадии имаго. У имаго жуков (Eleuterata) лабиальные щупики обычно 3-члениковые; у личинок жуков 3-члениковые, 2-члениковые или 1-члениковые. У веерокрылых нижняя губа и ее щупики утрачены (см. ниже, Strepsiptera). У имаго сетчатокрылообразных (Neuropteroidea) лабиальные щупики обычно 3-члениковые, редко 2-члениковые или 4-члениковые; у личинок сетчатокрылообразных лабиальные щупики либо 3-члениковые (рис. 10.5.3A, 10.6.4A и 10.6.6C), либо дополнительно расчленены (рис. 10.4.6A), либо утрачены. У имаго перепончатокрылых число члеников лабиального щупика, вероятно, исходно увеличено до 4; бывает от 1 до 4 члеников; у личинок перепончатокрылых лабиальный щупик 3-члениковый или одночлениковый. У имаго скорпионниц лабиальные щупики 2-члениковые, у блох бывают вторично расчленены или нерасчлененные (Aphaniptera); у личинок скорпионниц и блох лабиальные щупики 2-члениковые или вестигиальные. У имаго двукрылых лабиальные щупики преобразованы в нерасчлененные лабеллумы (см. ниже, Diptera); у личинок двукрылых нижняя губа сильно редуцирована и щупики не выражены. У имаго ручейников (Trichoptera) лабиальные щупики исходно 3-члениковые; у некоторых представителей модифицированы, с уменьшенным числом члеников; иногда вестигиальные или утрачены; у личинок лабиальные щупики утрачены. У имаго чешуекрылых (Lepidoptera) лабиальные щупики имеют от 1 до 4 члеников, у большинства 3-члениковые; у личинок лабиальные щупики 2-члениковые.
● Грудь. Туловище, т.е. отдел, следующий после головы (включающий сегменты начиная с V посторального), разделено на две тагмы – грудь, включающую первые 3 сегмента (V–VII посторальные) и брюшко, включающее остальные 10 сегментов. Локомоторные конечности (ноги) сохраняются только на грудных сегментах; конечности брюшных сегментов редуцированы или модифицированы так, что утратили функцию ног. Грудные сегменты отличаются от брюшных не только по строению и наличию конечностей, но и по расположению трахеостий.
Три сегмента груди называются соответственно переднегрудь (prothorax), среднегрудь (mesothorax) и заднегрудь (metathorax). Тергит переднегруди называется пронотум (pronotum), тергит среднегруди – мезонотум (mesonotum), тергит заднегруди – метанотум (metanotum); стерниты называются соответственно простернум (prosternum), мезостернум (mesosternum) и метастернум (metasternum), а плейральные области каждого из этих сегментов – соответственно проплевры (propleura), мезоплевры (mesopleura) и метаплевры (metapleura). Каждый из трех сегментов груди несет пару локомоторных конечностей, называемых соответственно передние ноги, средние ноги, и задние ноги.
Расположение трахеостий. Трахеостии (парные латеральные – см. ниже) по-разному расположены на груди и брюшке. В примитивном случае имеются следующие пары трахеостий (рис. 6.1.1 и 6.3.1A). Первая пара, стеноторакальная – между переднегрудью и среднегрудью (сохраняется у Diplura и Amyocerata); вторая пара, мезоторакальная – на среднегруди (сохраняется у Diplura и Protura); третья пара пара, криптоторакальная – между среднегрудью и заднегрудью (сохраняется у Diplura и Amyocerata); четвертая пара, метаторакальная – на заднегруди (сохраняется у Diplura и Protura) и V–XII пары, абдоминальные – на каждом из I–VIII сегментов брюшка (все или некоторые из них сохраняются у Diplura и Amyocerata). Таким образом, могут присутствовать 2 пары межсегментарных трахеостий груди (I и III пары), 2 пары сегментарных трахеостий груди (II и IV пары) и 8 пар сегментарных трахеостий брюшка (V–XII пары); все или часть из них могут служить дыхальцами.
У Amyocerata имеется максимум 2 пары межсегментарных дыхалец на груди и 8 пар сегментарных дыхалец на брюшке, иногда часть из них утрачена (см. ниже, Amyocerata). Среди Entognatha, у двухвосток имеется до 4 пар дыхалец на груди и до 7 пар дыхалец на брюшке (см. ниже, Diplura), иногда часть из них утрачена; у бессяжковых могут присутствовать 2 пары сегментарных дыхалец на груди (не гомологичных межсегментарным грудным дыхальцам Amyocerata), иногда они утрачены (см. ниже, Protura); у ногохвосток первичные трахеостии утрачены (см. ниже, Collembola).
● Строение ноги (рис. 6.1.2). Первый членик ноги (коксоподит – см. Euarthropoda), называемый здесь тазик, или кокса (coxa), на всех грудных ногах лишен гнатобазы. Обычно (но не всегда) тазик сочленяется с туловищем двумя мыщелками – дорсо-латеральным и вентро-медиальным. Дорсо-латеральный мыщелок соединяет тазик с плейритом. Вентро-медиальный мыщелок соединяет тазик либо со стернитом, либо с трохантином. Трохантин (trochantinus), если имеется, представляет собой более или менее обособленный участок плейрита, прилегающий к переднему краю основания тазика (рис. 7.1.1B).
К тазику посредством подвижного двухмыщелкового сочленения причленен сравнительно короткий вертлуг, или трохантер (trochanter), двигающийся дорсовентрально относительно тазика. К вертлугу малоподвижно причленено относительно длинное бедро, или фемур (femur), обычно имеющее то же направление, что и вертлуг. К бедру подвижно причленена голень, или тибиа (tibia); бедро и голень образуют единственное коленное сочленение, которое обычно является двухмыщелковым и позволяет голени совершать дорсовентральные движения в пределах от состояния вытянутого в одну линию с бедром до согнутого вентрально и прижатого к бедру (аналогично коленному суставу позвоночных). К голени причленена лапка, или тарсус (tarsus) (см. Atelocerata); у Entognatha лапка нерасчлененная, у Amyocerata она имеет до 5 члеников (см. ниже, Entognatha и Amyocerata). К лапке причленен претарсус (praetarsus), сгибающийся вентрально (см. Atelocerata) и несущий коготки (см. ниже).
Тазик (кокса), будучи коксоподитом, двигается мышцами, крепящимися в туловище; различные коксальные мышцы могут крепится к тергиту, плейриту, стерниту, мезодермальным сухожилиям и/или кутикулярным аподемам – к фурке, спине и/или плейральной аподеме (см. ниже, Pterygota). Вертлуг (трохантер), будучи первым члеником телоподита, двигается мышцами, крепящимися в тазике и в туловище; различные трохантеральные мышцы могут крепится к тергиту, плейриту, плейральной аподеме и/или фурке. Прочие членики ноги двигаются только мышцами, крепящимися в предыдущих члениках: к основанию бедра идут мышцы только из вертлуга; к основанию голени могут идти мышцы из вертлуга и бедра, к основанию лапки – из голени, к единственной аподеме претарсуса – из бедра и голени (рис. 6.1.2).
Других члеников ноги нет. Бывает лишь явно вторичное расчленение или слияние названных члеников.
● Коготки претарсуса. Претарсус (самый дистальный отдел ноги, сгибаемый унгуитрактором – см. Atelocerata) имеет два боковых коготка (ungues) – т.е. два коготка, причлененных по бокам исходного срединного коготка (дактилоподита); таким образом, претарсус исходно несет 3 коготка (рис. 6.1.2). Среди Entognatha дактилоподит и оба боковых коготка имеются у Diplura, тогда как у ногохвосток и протур боковые коготки утрачены (см. ниже, Ellipura). У Diplura оба боковых коготка причленены к основанию дактилоподита и двигаются вместе с ним (рис. 6.3.1D). Среди Amyocerata дактилоподит и оба боковых коготка имеются у большинства Zygentoma (рис. 6.7.3A), тогда как у Microcoryphia и у всех крылатых неасекомых (Pterygota) дактилоподит редуцирован, так что имеются лишь два боковых коготка. У Amyocerata все коготки (три коготка у Zygentoma или два коготка у Microcoryphia и Pterygota) вентрально сочленены с унгуитрактором (см. Atelocerata), а дорсально подвижно причленены к вершине последнего членика лапки (рис. 6.7.3A, 7.2.4D). Иногда имеется лишь один коготок, или все коготки утрачены. У многих Pterygota, помимо двух коготков, претарсус несет вторичные придатки того или иного строения, называемые аролий (arolium), эмподий (empodium), пульвилла (pulvillus) и др.
● I–IX сегменты брюшка и их придатки. В примитивном случае стернит каждого из первых девяти уромеров несет на заднем крае пару подвижных стерностилей, а между ними пару кинетапофизов (рис. 6.3.2A, 6.6.3).
Стерностиль (sternostylus), или стилус (styus) представляет собой подвижный одночлениковый придаток палочковидной формы. К основанию стерностиля подходит мышца, неподвижный конец которой крепится к стерниту или к стилигеру (см. ниже). Внутри самого стерностиля мышц никогда не бывает. Стерностиль может нести неподвижный склеротизованный заостренный апикальный шип.
Кинетапофиз (kinetapophysis) представляет собой либо мембранозный втяжной пузырек (vesicula retractilis), способный вворачиваться и втягиваться в тело, либо невтягивающийся подвижный отросток. К основанию кинетапофиза, как и к основанию стерностиля, подходит мышца, неподвижный конец которой крепится к стерниту или к стилигеру.
Стернит может быть цельным, либо разделен поперечным стилигеральным швом на переднюю часть – престилигер (praestyliger) и заднюю часть – стилигер (styliger). В последнем случае все мышцы, идущие к стерностилям и кинетапофизам крепятся в пределах стилигера. Стилигер может выступать назад в виде пластины или пары пластин, на заднем крае которых находятся стерностили и кинетапофизы. Единый непарный стилигер, несущий левые и правые стерностили и/или кинетапофизы, можно называть бистилигером (bistyliger), а обособленные левый или правый стилигеры, каждый из которых несет один стерностиль и/или один кинетапофиз, можно называть унистилигером (unistyliger).
На прегенитальных сегментах брюшка (т.е. на I–VII уромерах) стерностили и кинетапофизы развиты только у первичнобескрылых насекомых – Triplura и Diplura. Кинетапофизы этих сегментов обычно представляют собой втяжные пузырьки, лишь у части Diplura кинетапофизы I уромера являются невтягивающимися придатками. У Pterygota стерностили и кинетапофизы всех прегенитальных сегментов утрачены.
На генитальных сегментах брюшка (т.е. на VIII–IX уромерах) у самки кинетапофизы либо представляют собой створки яйцеклада (см. Amyocerata), либо утрачены. У Triplura стерностили генитальных сегментов сходны со стерностилями прегенитальных сегментов; у крылатых насекомых стерностили сохраняются только на IX уромере. У самцов поденок стерностили IX уромера сохраняют подвижность и преобразованы во вторичночленистые половые придатки (см. ниже, Ephemeroptera); у всех прочих крылатых насекомых утрачены мышцы, подходящие к стерностилям (см. ниже, Metapterygota). У многих крылатых насекомых утрачены все производные стерностилей и кинетапофизов.
Предполагают, что унистилигеры, стерностили и кинетапофизы являются вестигиями утраченных абдоминальных ног, но их гомология с частями ноги неясна. Стилигеры часто называют «кокситами», что связано с предположением об их гомологии с коксам ног. Однако, в отличие от кокс, стилигеры являются не обособленными члениками, а составляют часть вентральной стенки тела; они всегда расположены не по бокам, а позади стернита; стилигеры неподвижны, и мышцы из туловища к ним не подходят.
На основании того, что строение придатков брюшка имеет явное сходство у Diplura (относящихся к Entognatha) и у Triplura (относящихся к Amyocerata), можно заключить, что это строение является исходным для Hexapoda.
● Десятый сегмент брюшка. Брюшко исходно состоит из 10 сегментов (уромеров). Десятый уромер, на котором располагается анальное отверстие, не имеет структур, присущих первым девяти уромерам. Среди Entognatha десятый уромер имеется только у Diplura, тогда как у ногохвосток и протур он полностью утрачен (см. ниже, Ellipura). Среди Amyocerata десятый уромер имеется у всех представителей, но у некоторых он упрощен и/или уменьшен. Десятый уромер может нести парные подвижные придатки, называемые «церками», но имеющие принципиально разное строение у Diplura и у Amyocerata (см. ниже, Diplura и Amyocerata). Принято считать, что в состав брюшка исходно вошло не 10, а 11 сегментов, но эта интерпретация строения брюшка ошибочна (Клюге 2020).
● Трахейная система. Трахейная система (возникавшая у разных членистоногих независимо несколько раз – см. Gnathopoda), у Hexapoda имеет единое происхождение, о чем свидетельствует единое для Hexapoda расположение парных сегментарных и межсегментарных трахеостий (см. выше). Обычно трахеостия находится между тергитом и стернитом – либо на плейральной мембране, либо на плейральном склерите (это может быть цельный плейрит или один из мелких склеритов). Иногда, благодаря разрастанию тергита или стернита на бока тела, трахеостии оказываются расположенными не между тергитом и стернитом, а на тергите (рис. 10.7.1) или на стерните (рис. 9.8.2A). Иногда расположение трахеостий не вполне соответствует внешней сегментации туловища: межсегментарная трахеостия может располагаться не только на межсегментной мембране, но и на склерите впереди лежащего сегмента или на склерите позади лежащего сегмента; cегментарная трахеостия может быть смещена на межсегментную мембрану или даже на склерит соседнего сегмента.
Тогда как расположение трахеостий едино для Hexapoda, расположение отходящих от них трахей довольно разнообразно. Трахеи ветвятся. Пучки трахей, отходящих от разных трахеостий, могут быть независимыми, либо анастомозируют. Во многих случаях трахеи, идущие от всех трахеостий одной стороны тела, соединены общим продольным латеральным трахейным стволом; такой ствол имеется у части Diplura, Zygentoma и многих Pterygota. Кроме пары латеральных стволов, у некоторых насекомых имеются другие анастомозы, соединяющие трахеи разных трахеостий; иногда они образуют дополнительные парные продольные стволы – дорсальные, вентральные и/или висцеральные; иногда имеются поперечные вентральные и/или дорсальные анастомозы, соединяющие трахейные стволы левой и правой половины тела. Возможно, пара латеральных стволов является исходной для Hexapoda, а прочие стволы и анастомозы возникли независимо в разных группах. При линьке старая кутикула, выстилающая трахейный ствол, рвется на фрагменты по числу трахеостий, с которыми она связана, и каждый его фрагмент вместе со старой кутикулой трахей, отходящих от этого участка ствола, выходит через свою трахеостию.
Среди Hexapoda только у Collembola трахейная система не соответствует вышеописанной; судя по всему, у ногохвосток трахейная система исходно была утрачена, после чего заново возникла трахейная система иного строения, с дыхальцами на шее (см. ниже, Collembola).
В большинстве случаев трахеостия представляет собой дыхальце – т.е. открываются наружу отверстием, через которое воздух попадает в трахеи; дыхальца могут иметь простое строение, либо снабжены фильтрующими и/или замыкательными аппаратами того или иного строения.
Иногда трахеостии определенных пар замкнуты и не функционируют. Нефункционирующая трахеостия снаружи может быть не выражена, но внутри от кутикулы отходит трахея (которая закладывается как впячивание кутикулы); при линьке старая кутикулярная выстилка трахеи выходит через открытую трахеостию в новой неокрепшей кутикуле, после чего трахеостия замыкается и новая кутикула затвердевает так, что трахеостия остается постоянно замкнутой и не функционирует. Такие нефункционирующие трахеостии свойственны, в частности, некоторым личинкам, специализированным к обитанию в воде; в результате метаморфоза на месте нефункционирующих трахеостий появляются функционирующие дыхальца; таким образом, в процессе онтогенеза сохраняется один и тот же набор трахеостий, но может меняться их способность функционировать как дыхальца. Реже трахеостии определенных пар утрачены.
У некоторых насекомых имеются вторичные дыхальца; в этом случае дыхальце находится не на самой трахеостии, а рядом с ней, и идущая к нему трахея отходит от трахеи, идущей от трахеостии; при этом трахеостия может быть замкнутой; при линьке старая кутикулярная выстилка всех трахей, включая дыхальцевую трахею, выходит не через вторичное дыхальце, а через трахеостию. Вторичные дыхальца имеются у многих личинок двукрылых.
Некоторые насекомые вторично перешли к водному образу жизни и приспособились дышать растворенным в воде кислородом. В этом случае трахейная система сохраняется, так что дыхание осуществляется парадоксальным образом: растворенный в окружающей воде кислород через покровы насекомого диффундирует в трахеи, переходя в газообразное состояние, по трахеям разносится по телу, а в тканях опять переходит в водный раствор. У некоторых таких водных насекомых диффузия кислорода через покровы в трахеи происходит преимущественно в выступающих тонкостенных придатках – трахейных жабрах (branchiae tracheales); в разных таксонах в качестве трахейных жабр служат те или иные придатки – максиллярные и/или лабиальные щупики, тергалии (см. ниже, Ephemeroptera), эпипрокт и/или парапрокты (см. ниже, Zygoptera), либо новообразования, независимо возникшие у разных насекомых на тех или иных частях тела. У многих из этих водных насекомых трахейная система замкнутая, то есть трахеостии постоянно закрыты и не служат дыхальцами; в некоторых случаях одновременно имеются и трахейные жабры, и функционирующие дыхальца. У некоторых водных насекомых брюшко несет латеральные отростки, которые можно назвать придыхальцевыми трахейными жабрами: трахея, входящая в такой отросток, начинается непосредственно от трахеи, идущей от дыхальца того же сегмента брюшка (подобно трахеи, идущей к вторичному дыхальцу – см. выше); сходные по строению придыхальцевые трахейные жабры независимо друг от друга возникли у некоторых личинок крылатых насекомых (рис. 7.3.1A, 10.6.2A, 10.6.6A, 10.15.1C).
Признак Hexapoda неясного филогенетического значения
● Форма линочного шва. Линочный шов, по которому при линьке трескается старая кутикула (см. Gnathopoda), обычно имеет следующую форму (рис. 7.2.3С): на головной капсуле имеется пара лобных (или фронтальных) швов (suturae frontales), которые кзади сходятся в один непарный продольный дорсальный шов, идущий далее назад медиально по голове и по тергитам груди. Область головы впереди и медиальнее лобных швов принято называть лоб (frons). Такая форма линочного шва свойственна подавляющему большинству преимагинальных возрастов насекомых; лишь в немногих случаях форма линочного шва изменена.
Геологический возраст. Девон – ныне (о древнейшем известном представителе – см. ниже, Collembola).
Статус Hexapoda. В некоторых классификациях таксон Hexapoda отсутствует. В литературе фигурирует утверждение, будто бы существует обоснованная и широко признанная теория о полифилетическом происхождении Hexapoda. В действительности никто никогда не выдвигал и не обосновывал теорию о полифилии Hexapoda, а недоразумение вызвано тем, что нетипифицированное название Insecta используется в качестве рангового (см. раздел I.6.6.2). Обоснованием теории о полифилии Hexapoda могли бы быть какие-нибудь признаки, считающиеся апоморфиями и являющиеся общими для какой-либо отдельной группы внутри Hexapoda и какой-либо группы животных, не входящих в Hexapoda (например, каких-либо Myriapoda). Однако такие апоморфии не описаны.
Классификации Hexapoda.
I. Деление Hexapoda на Apterygota и Pterygota. Долгое время было принято деление Hexapoda на первичнобескрылых (Apterygota) и крылатых (Pterygota). Таксон Apterygota Lang 1888 – первичнобескрылые {syn.circ.: Thysanoura Latreille 1796 = Seticaudes Cuvier 1805 = Nematoura Dumeril 1805 = Thysanura Leach 1815a = Thysanuria Rafinesque 1815 = Monomorphes Lucas 1840 = Apterygogenea Brauer 1885 = Apterota Haeckel 1896 = Aptilota Lameere 1935 = Epimetabola Weber 1933 (название для типа метаморфоза)} объединяет Entognatha и Triplura. Он является парафилетическим и предковым для голофилетического таксона Pterygota.
II. Деление Hexapoda на Entognatha и Amyocerata. Эта классификация основана на представлении о том, что Triplura имеют общего предка с Pterygota. На основании этого Triplura и Pterygota объединяются в голофилетический таксон Amyocerata, тогда как Diplura, Collembola и Protura объединяются в другой голофилетический таксон – Entognatha (об апоморфиях Entognatha и Amyocerata см. ниже). При этом таксон Apterygota не признается, поскольку является парафилетическим; таксон Pterygota в этой классификации оказывается подчиненным в составе Amyocerata. В данной книге мы придерживаемся именно этой классификации:
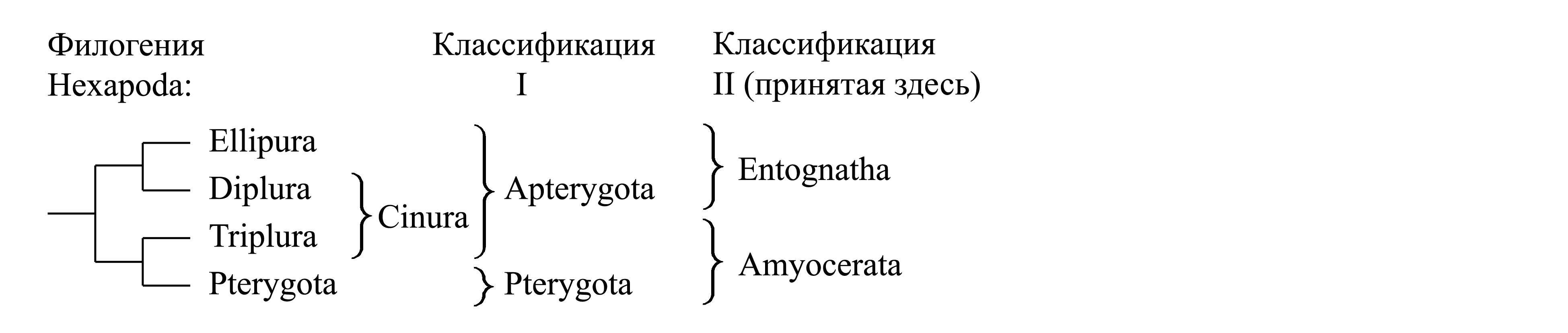
Здесь мы принимаем деление Hexapoda на Entognatha и Amyocerata.
Hexapoda – шестиногие насекомые
![]() 1. Entognatha
– скрыточелюстные
1. Entognatha
– скрыточелюстные
![]() 1.1. Diplura
– двухвостки
1.1. Diplura
– двухвостки
![]() 1.2. Ellipura
1.2. Ellipura
![]() 1.2.1. Collembola
– ногохвостки
1.2.1. Collembola
– ногохвостки
![]() 1.2.2. Protura
– бессяжковые
1.2.2. Protura
– бессяжковые
![]() 2. Amyocerata
– мезмышцеусые
2. Amyocerata
– мезмышцеусые
![]() 2.1. Triplura
– трехвостки
2.1. Triplura
– трехвостки
![]() 2.1.1. Zygentoma
2.1.1. Zygentoma
![]() 2.1.2. Microcoryphia
2.1.2. Microcoryphia
![]() 2.2. Pterygota
– крылатые наскомые . . .
2.2. Pterygota
– крылатые наскомые . . .
Подробнее см. Н.Ю. Клюге 2020. «Систематика насекомых и принципы кладоэндезиса».