КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ

![]()
|
КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IX-1.2.2.2. Arthroidignatha Spinola 1850 – членистохоботные
(Somobialia
Eucaryota Metazoa Monostomata
Haemataria Peritoneata
Metameria
Gnathopoda Euarthropoda
Mandibulata Atelocerata
Hexapoda Amyocerata
Pterygota
Metapterygota Neoptera
Eumetabola
Parametabola
Acercaria
Condylognatha Arthroidignatha)
(рис. 9.6.1–9.12.4)
(рис. 9.6.1, 9.7.1,
9.7.2)
|
Названия, однозначно соответствующие по циркумскрипции: — Semivaginata Schluga 1767; — Arthroidignatha Spinola 1850; NomCirc. — Hemipteroidea Handlirsch 1903; — Hemipteradelphia = Panhemiptera Crampton 1924; — Hemipteriforma Metcalf 1951; — Hemipteroida Müller 1963; — Hemipterida Boudreaux 1979b; — Homopteroidea Brodsky 1994. По циркумскрипции также соответствует: — «Hemiptera» sensu Geoffroy 1762 (non Linnaeus 1758); — «Ryngota» auct. (non Fabricius 1775); — «Rhyngota» auct. (non Fabricius 1803); — «Rhynchota» auct. (non Burmeister 1835; nec Rhynchota Billberg 1820); — таксонам под типифицированными названиями, образованными от родового названия Cimex: Cimicida, Cimicidea и др. Иерархическое название: Cimex/f2=Cicada/g4 [f:1781(ord.3); g:L.1758(gen.195)] (sine Thrips; incl. Aphis). |
|
Ранг. Обычно отряд или надотряд.
Названия. Иногда этому таксону приписывают название Ryngota Fabricius 1775 или его измененное написание Rhyngota Fabricius 1799. В действительности оба эти названия не соответствуют по своей исходной циркумскрипции рассматриваемому таксону, так как Фабрициус относил к Ryngota, помимо прочего, роды Pulex и Thrips (т.е. таксоны, называемые ныне Aphaniptera и Thysanoptera).
Широко признанным названием этого таксона является Rhynchota (измененное Rhyngota), однако и оно не соответствует ему по исходной циркумскрипции, поскольку исходно название Rhynchota Burmeister 1835 относилось к таксону, включающему, помимо прочего, вшей (см. Siphunculata).
Часто этот таксон называют также Hemiptera. Старинное название Hemiptera в XVIII веке давалось самым различным таксонам: сборной группе, включавшей скорпионов, клопов, светляков, прямокрылых и муравьев (Linnaeus 1735); группе, объединяющей насекомых с умеренно жесткими передними крыльями – хоботных и прямокрылых в широком смысле (Linnaeus 1789), только прямокрылых в широком смысле (Retzius 1783) или только хоботных в широком смысле. В 10-м издании «Systema Naturae» (Linnaeus 1758) (являющемся исходным для приоритета зоологических названий – см. I.6.3.1.2) название Hemiptera отнесено к группе, включавшей не только членистохоботных, но и трипсов, т.е. по циркумскрипции соответствующей Condylognatha. В таком значении это название широко употреблялось в течение всего XIX века; некоторые авторы середины и конца XIX века относили к Hemiptera, помимо членистохоботных и трипсов, также вшей. В XX веке название Hemiptera в этих значениях почти не используется, и чаще всего употребляется в одном из двух сильно различающихся значений: либо для Arthroidignatha, либо для Heteroptera. Таким образом, во избежание путаницы желательно вообще не использовать название «Hemiptera».
Название Arthroidignatha Spinola 1850 однозначно соответствует по циркумскрипции данному таксону; оно было предложено автором, установившем этот таксон и давшем ему правильную характеристику.
Более раннее забытое название Semivaginata Schluga 1767 является латинским эквивалентом греческого названия «Hemelytrata», т.е. отражает признак, свойственный лишь подчиненному таксону (см. ниже, Hemelytrata); употребление этого названия может вызвать такие же недоразумения, как употребление сходного по значению названия «Hemiptera».
Аутапоморфии Arthroidignatha
Колюще-сосущий ротовой аппарат (см. Condylognatha) усложнен (рис. 9.6.1, 9.7.1A, 9.7.2): в состав внутреннего хоботка (proboscis interior), помимо стилетов максиллярных лациний, видоизмененных особым образом, вошли видоизмененные мандибулярные стилеты; внутренний хоботок заключен во внешний хоботок (proboscis exterior), образованный сильно видоизмененной нижней губой; слюнной шприц имеет особое строение (см. ниже). В таком аппарате весь внутренний хоботок работает как единая конструкция; он в совершенстве приспособлен для прокалывания покровов и высасывания жидкой пищи, которой исходно является сок растений. Со строением ротового аппарата связаны общие особенности эволюции, проявляющиеся в разных таксонах членистохоботных (см. ниже).
Поскольку стилеты внутреннего хоботка скрыты внутри внешнего хоботка, а щупики утрачены, снаружи весь ротовой аппарат выглядит как цельный непарный членистый внешний хоботок (с чем связано название Arthroidignatha); лишь спереди к основанию хоботка прижата более короткая верхняя губа.
Ротовой аппарат принципиально одинаково устроен у имаго и у личинок всех возрастов; лишь у непитающихся стадий ротовой аппарат может быть вестигиальным или утрачен.
Ниже, как и в характеристике Condylognatha, ротовые части условно описаны так, как они расположены при гипогнатном положении головы (когда ротовые придатки направлены вершинами вентрально, клипеус и верхняя губа находятся спереди, а нижняя губа сзади), хотя среди Arthroidignatha имеются также представители с опистогнатной и прогнатной головой.
● Модификация мандибул (рис. 9.6.1). Мандибула (скрытая в кармане и дистально вытянутая в стилет – см. Condylognatha) приобрела подвижный мандибулярный рычаг и мышцы-протракторы, идущие к особой мандибулярной пластинке головы, а мандибулярные стилеты сцеплены с максиллярными, образуя вместе с ними единый внутренний хоботок.
Из двух мыщелков мандибулы сохраняется только один – исходно задне-латеральный. Имеется мандибулярный рычаг, который представляет собой обособленный палочковидный склеротизованный участок мандибулы, исходно являющийся проксимальным и несущий мыщелок. От мыщелка, соединяющего мандибулярный рычаг с краем головной капсулы, мандибулярный рычаг тянется перпендикулярно поверхности головной капсулы вглубь мандибулярного футляра и там сочленен с остальной частью мандибулы, которая представляет собой мандибулярный стилет. Рычаг и стилет мандибулы, подвижно соединенные под углом друг к другу, по строению и принципу действия напоминают рычаг и стилет максиллярной лацинии, хотя их происхождение совершенно различно (максиллярный рычаг, в отличие от мандибулярного, является внутренней аподемой – см. Condylognatha). Подвижно сочлененный рычаг позволяет мандибуле вдвигаться и выдвигаться, совершая прокалывающие движения.
Благодаря такому строению мандибулы ее задняя мышца (исходно являющиеся аддуктором – рис. 4.6A:PDm) служат ретрактором (рис. 9.6.1A:cr-md, tnt-md). Латеральные области головной капсулы, которые исходно вмещают неподвижные основание передних мышц мандибул, сместилась вентрально и находятся по бокам от клипеуса; эти участки головной капсулы называются мандибулярные пластинки, или лорумами (lora). В результате такого смещения передняя мышца мандибулы (исходно являющаяся абдуктором – рис. 4.6A:ADm) служит протрактором мандибулы (рис. 9.6.1A:lor-md). Задний край мандибулярной пластинки плотно смыкается с передним краем максиллярной пластинки (т.е. стипесом максиллы); между ними имеется щель, ведущая в мандибулярный футляр (исходно эта щель разделяет клипеус и максиллярную пластинку – см. Condylognatha).
Мандибулярный стилет имеет желоб на внутренней (медиальной) стороне, благодаря чему пара мандибулярных стилетов плотно охватывает с боков трубку, образованную парой максиллярных стилетов (см. ниже); в результате этого мандибулярные и максиллярные стилеты образуют единый внутренний хоботок. Двигаясь поочередно, мандибулярные стилеты прокалывают субстрат. У многих Heteropteroidea мандибулярные стилеты не только охватывают максиллярные, но и прочно сцепляются с ними с помощью продольных гребней и пазов, образующих скользящие соединения.
● Модификация максилл (рис. 9.6.1, 9.7.2D). Наружные части максилл (не имеющих разделения на кардо и стипес – см. Acercaria) в основании слиты с головной капсулой и представляют собой пару максиллярных пластинок – неподвижных продолжений латеральных стенок головной капсулы. Соответственно, максиллярная пластинка служит местом прикрепления мышц, выдвигающие максиллярный стилет; к максиллярной пластинке или к отходящей от нее аподеме крепится максиллярный рычаг (см. Condylognatha). Максиллярная пластинка отделена от лежащей впереди нее мандибулярной пластинки щелью, ведущей в карман, в котором расположены максиллярный и мандибулярный стилеты. Максиллярный щупик полностью утрачен, так что снаружи максиллярные пластинки утратили сходство с максиллами и кажутся участками головной капсулы.
Максиллярные стилеты (лацинии, исходно образующие канал для проведения пищи и слюны – см. Condylognatha) имеют такую форму поперечного сечения, что, складываясь вместе, они вместо одного канала образуют два изолированных канала, по которым жидкости движутся в противоположных направлениях: передний (дорсальный) канал служит для проведения жидкой пищи в глотку; задний (вентральный) канал служит для проведения слюны, которая подается в него из отростка гипофаринкса (см. ниже) (рис. 9.6.1B, 9.7.2C). Максиллярные стилеты входят в состав внутреннего хоботка вместе с охватывающими их мандибулярными стилетами (см. выше).
● Модификация гипофаринкса (рис. 9.7.2C–D). Гипофаринкс образует слюнной шприц, усложненный по сравнению с подобным образованием у Thysanoptera (см. Condylognatha); этот шприц накачивает слюну в задний из двух каналов, образованных максиллярными стилетами. Латеральные края гипофаринкса (завернутые назад и обрамляющие слюнной желоб – см. Condylognatha) смыкаются сзади (вентрально) по средней линии так, что образуют непарный канал шприца; в глубине этого канала имеется расширение – камера шприца, в которую сзади открывается непарный проток слюнных желез. Камера имеет склеротизованные стенки, а ее проксимальная стенка образует поршень – обособленный склеротизованный выступ, плотно входящий в камеру. Мышцы гипофаринкса сильно увеличены и крепятся к непарному сухожилию, отходящему проксимально от поршня шприца. При сокращении мышц поршень шприца отводится, камера расширяется, и слюна накачивается из слюнного протока в камеру; при расслаблении мышц поршень под действием упругости стенок камеры вдвигается в камеру, и слюна под давлением накачивается в слюнной канал, по которому идет до вершины гипофаринкса. Вершина гипофаринкса вставлена между максиллярными стилетами так, что входит в задний канал, образованный ими (рис. 9.7.2C).
● Модификация нижней губы (рис. 9.6.1, 9.7.2B). Нижняя губа (исходно упрощенная – см. Acercaria) крайне видоизменена и превращена в членистый внешний хоботок (proboscis exterior): она сильно удлинена, ее латеральные края завернуты вперед и смыкаются (но не срастаются) спереди по прямой продольной линии так, что вся нижняя губа образует трубку, окружающую внутренний хоботок; эта трубка подразделена поперечными швами на несколько подвижных члеников.
Судя по всему, исходным для Arthroidignatha является 3-члениковый внешний хоботок. Внешний хоботок имеет 3 членика у Aleyroptera (рис. 9.10.1A), у части Gallinsecta, у всех Auchenorrhyncha-Euhomoptera (рис. 9.7.2A–B), у примитивных Auchenorrhyncha-Subtericornes и у Coleorrhyncha (рис. 9.8.1D). У некоторых Auchenorrhyncha-Subtericornes внешний хоботок 4-члениковый или 5-члениковый благодаря разделению исходного второго членика. У большинства клопов хоботок 4-члениковый благодаря наличию дополнительного членика, находящегося проксимальнее входа в базальную аподему (см. ниже, Heteroptera и рис. 9.8.2A). У тлей хоботок 4-члениковый благодаря разделению на два исходного третьего членика (рис. 9.11.2). В некоторых случаях число члеников хоботка вторично уменьшено из-за слияния тех или иных члеников, иногда расчленение хоботка совсем утрачено. У листоблошек хоботок 2-члениковый (см. ниже, Saltipedes и рис. 9.9.1A, 9.9.2A). У некоторых Gallinsecta число члеников хоботка сокращено от трех до 2 или 1. У некоторых клопов число члеников сокращено от исходных четырех до 3 или 1.
Во внешнем хоботке отсутствуют парные придатки, характерные для нижней губы других насекомых – глоссы, параглоссы и лабиальные щупики; так что неясно, каким частям исходной нижней губы гомологичны те или иные части внешнего хоботка.
Функционирование хоботка. Исходно хоботок приспособлен для питания соками и клеточным содержимым сосудистых растений: фитофагия свойственна всем Plantisuga, Auchenorrhyncha, Coleorrhyncha и части Heteroptera; лишь среди Heteroptera, помимо фитофагов, имеются факультативные и облигатные хищники, высасывающие гемолимфу из пойманных насекомых; несколько видов Heteroptera приспособились к питанию кровью позвоночных животных.
Все функции хоботка – и прокалывание покровов, и проведение жидкой пищи из субстрата в глотку, и проведение слюны из саливария в субстрат – осуществляются внутренним хоботком, который состоит из сложенных вместе или сцепленных парных мандибулярных и максиллярных стилетов. Мандибулярные стилеты, двигаясь поочередно, проникают в субстрат, а пара сомкнутых максиллярных стилетов, образующих пищевой и слюнной каналы, вдвигается в канал, образованный сомкнутыми мандибулярными стилетами.
Внутренний хоботок проходит внутри желоба внешнего хоботка, образованного нижней губой. Спереди к основанию внешнего хоботка прижата более короткая верхняя губа; иногда внутренний хоботок проходит в желобе на задней стороне верхней губы, а дистальнее входит в желоб на передней стороне внешнего хоботка (рис. 9.11.2). В покое стилеты внутреннего хоботка не выступают за вершину внешнего хоботка и не видны снаружи; при питании насекомое приставляет вершину внешнего хоботка к субстрату, затем стилеты, скользя друг вдоль друга, по очереди выдвигается из вершины внешнего хоботка, и, благодаря этому, внутренний хоботок постепенно проникает в субстрат.
Внутренний хоботок может быть умеренной длины, так что во втянутом состоянии он остается прямым и помещается в голове и внешнем хоботке. Но у некоторых фитофагов, имеющих мелкие размеры и питающихся соками из глубоко лежащих тканей растения, внутренний хоботок удлинен настолько, что во втянутом состоянии образует петли той или иной формы, которые помещаются либо снаружи головы (у личинок Saltipedes), либо внутри кармана того или иного строения, называемого крумена (crumena) (у имаго Saltipedes, личинок и самок Gallinsecta, личинок Aleyroptera и у некоторых Heteroptera). Внутренний хоботок как единое целое образует петли, а составляющие его четыре стилета могут скользить друг вдоль друга, не раздвигаясь. Поэтому, когда внутренний хоботок сложен в виде петли, его выдвигание осуществляется следующим образом: сначала, благодаря поочередному сокращению протракторов и скольжению стилетов по петле, стилеты поочередно выдвигаются, но петля сохраняет свою форму и размер; затем весь внутренний хоботок зажимается в желобе внешнего хоботка дистальнее петли, а ретракторы стилетов, сокращаясь все одновременно, втягивают основание внутреннего хоботка и тем самым уменьшают изгиб его петли; затем цикл повторяется, пока петля не выпрямится полностью (Weber 1933).
Особенности эволюции Arthroidignatha, связанные с наличием хоботка. В связи с приобретением колюще-сосущего хоботка, в совершенстве приспособленного для сосания сока из живых растений, у Arthroidignatha появились новые возможности в эволюции образа жизни. В то время как большинство наземных животных для добывания пищи вынуждены постоянно перемещаться, сосущие фитофаги могут добывать пищу, не перемещаясь. Членистохоботное небольших размеров может, внедрив свой внутренний хоботок в ткани растения, достичь им проводящего сосуда так, чтобы поступающего по сосуду сока было достаточно для полного обеспечения насекомого пищей. В этом случае насекомому выгоднее оставаться на одном месте и питаться из одного и того же сосуда, чем перемещаться и искать подходящий сосуд заново. В связи с этим среди членистохоботных, помимо обычных активно перемещающихся форм (Heteropteroidea, многие Auchenorrhyncha и др.) имеются малоподвижные и совсем неподвижные формы (личинки Aleyroptera, личинки и самки многих Gallinsecta и др.).
Другой особенностью эволюции членистохоботных является то, что у некоторых из них кишечник приобрел фильтрационную камеру того или иного строения, благодаря которой кишечник эффективно всасывает белки, которые в малом количестве содержатся в соке растения, а излишки воды с углеводами выходят из анального отверстия. Эти сахаристые экскременты, известные как падь, или медвяная роса, могут загрязнять растения, приводя к их угнетению и тем самым лишая насекомое возможности питаться, оставаясь на одном месте. Большую помощь малоподвижным и неподвижным членистохоботным в избавлении от жидких сахаристых экскрементов оказывают насекомые, поедающие эти экскременты. Это преимущественно имаго различных насекомых с полным превращением – двукрылые (Diptera), ручейники (Trichoptera), бабочки (Lepidoptera), перепончатокрылые (Hymenoptera) и др. В их питании белки не обязательны (поскольку имаго не растут – см. Pterygota), а углеводы важны для пополнения энергетических затрат. В некоторых случаях симбиоз Arthroidignatha с имаго Metabola привел к появлению особых коадаптаций; в частности, многие Plantisuga и некоторые личинки Auchenorrhyncha имеют поведенческие и морфологические приспособления к кормлению своими экскрементами рабочих муравьев.
Классификации Arthroidignatha
I. Деление Arthroidignatha на Homopterida и Heteroptera. В этой классификации все Arthroidignatha, не относящиеся к голофилетическому таксону Heteroptera, объединяются в парафилетический таксон Homopterida Pearce 1936, обычно неверно называемый «Homoptera» (исходно таксон Homoptera Latreille 1810 включал, помимо прочего, трипсов). Homopterida отличаются от Heteroptera следующими плезиоморфиями.
(1) Внешний хоботок отходит от задней части головы, позади него нет склеротизованной области (рис. 9.7.1A, 9.10.1A, 9.11.2A). Очевидно, это плезиоморфия, поскольку внешний хоботок образован нижней губой, которая исходно является слитыми конечностями последнего сегмента головы и своим основанием граничит непосредственно с шейной мембраной.
(2) Передние крылья не разделены на кориум и перепоночку (как у всех прочих насекомых кроме Heteroptera).
II. Деление Arthroidignatha на Hemelytrata и Plantisuga. В то время как таксон Homopterida характеризуется лишь плезиоморфиями, таксоны Hemelytrata и Plantisuga имеют апоморфии, свидетельствующие об их голофилии (см. ниже).
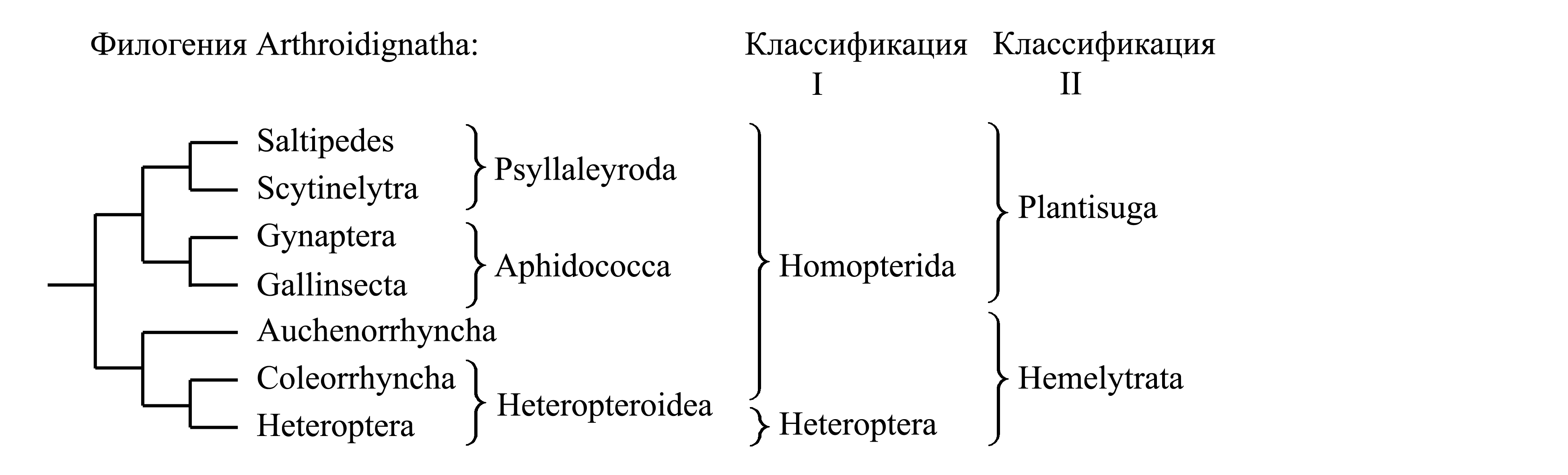
Здесь мы принимаем деление Arthroidignatha на Hemelytrata и Plantisuga.
Arthroidignatha – членистохоботные
![]() 1. Hemelytrata
1. Hemelytrata
![]() 1.1. Auchenorrhyncha – цикадообразные
1.1. Auchenorrhyncha – цикадообразные
![]() 1.2. Heteropteroidea
1.2. Heteropteroidea
![]() 1.2.1. Coleorrhyncha – пелоридииды
1.2.1. Coleorrhyncha – пелоридииды
![]() 1.2.2. Heteroptera – клопы
1.2.2. Heteroptera – клопы
![]() 2. Plantisuga
2. Plantisuga
![]() 2.1. Psyllaleyroda
2.1. Psyllaleyroda
![]() 2.1.1. Saltipedes (или Psyllida) – листоблошки
2.1.1. Saltipedes (или Psyllida) – листоблошки
![]() 2.1.2. Aleyroptera (или Aleyrodida) – белокрылки
2.1.2. Aleyroptera (или Aleyrodida) – белокрылки
![]() 2.2. Aphidococca
2.2. Aphidococca
![]() 2.2.1. Gynaptera (или Aphidida) – тли
2.2.1. Gynaptera (или Aphidida) – тли
![]() 2.2.2. Gallinsecta (или Coccida) – червецы и щитовки
2.2.2. Gallinsecta (или Coccida) – червецы и щитовки
Подробнее см. Н.Ю. Клюге 2020. «Систематика насекомых и принципы кладоэндезиса».