КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ

![]()
|
КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
X-1.3.3. Diptera Linnaeus 1758 – двукрылые
(Somobialia
Eucaryota Metazoa Monostomata
Haemataria Peritoneata
Metameria
Gnathopoda Euarthropoda
Mandibulata Atelocerata
Hexapoda Amyocerata
Pterygota
Metapterygota Neoptera
Eumetabola
Metabola Mecopteriformia Diptera)
(рис. 10.14.1–10.14.5)
(рис. 10.14.1,
10.14.2, 10.14.3,
10.14.4, 10.14.5)
|
Названия, однозначно соответствующие по циркумскрипции: — Diptera (Aristoteles) Linnaeus 1758; NomCirc. — Halterata Scopoli 1763; — Bialata Schluga 1767 — Halteriptera Clairville 1798; — Dipterae Robineau-Desvoidy 1830; — Antliognatha Spinola 1850; — Dipterida Pearce 1936. По циркумскрипции также соответствует: — «Antliata» sensu Burmeister 1835 (non Fabricius 1775); — таксонам под типифицированными названиями, образованными от родового названия Musca: Muscoides, Muscida. Иерархическое название: Musca/f1=Oestrus/g1 [f:1781(ord.7); g:L.1758(gen.220)] (incl. Tipula). |
|
Ранг. Обычно отряд.
Аутапоморфии Diptera
● Имагинальный ротовой аппарат и питание. У имаго ротовой аппарат преобразован в два хоботка, выполняющих две разные функции (рис. 10.14.2): (A) первичный кровососательный хоботок, образованный видоизмененными верхней губой, мандибулами, гипофаринксом и максиллярными лациниями, исходно служит для прокалывания кожи позвоночных и всасывания крови (см. ниже); он развит только у самок и используется ими после оплодотворения для обеспечения белком развивающихся яиц; (B) лижущий хоботок, образованный видоизмененными щупиками нижней губы (см. ниже), исходно служит для собирания жидкой углеводной пищи и используется обоими полами на всех этапах имагинальной жизни для восполнения энергетических затрат; жидкая пища, собираемая лижущим хоботком, сохраняется в особом вентральном дивертикуле кишки (см. ниже), из которого по мере надобности перекачивается в желудок; в отличие от нее, кровь, всасываемая первичным кровососательным хоботком, сразу попадает в желудок и переваривается; исходно у самок имеется гонотрофический цикл, благодаря которому потребляемая кровь эффективно используется для развития яиц (см. ниже). У самца кровососательного хоботка нет, а соответствующие части ротового аппарата либо отсутствуют, либо более или менее сходны с таковыми самки, но неспособны функционировать. Большинство кровососущих видов не имеет ясно выраженной специализации и способны питаться кровью разнообразных наземных позвоночных – лягушек, ящериц, птиц и млекопитающих, а также выползающих на сушу рыб-прыгунов и др.; некоторые виды кровососов более узко специализированы; некоторые двукрылые используют свой кровососательный ротовой аппарат для питания гемолимфой насекомых. При утрате способности к кровососанию мандибулы, как правило, становятся нефункционирующими или исчезают; это свойственно большинству двукрылых.
Как и у большинства других Metabola, у Diptera полностью утрачена эволюционная корреляция между строением имагинального и личиночного ротовых аппаратов. В отличие от имаго, у личинки ротовой аппарат либо сохраняет исходную грызущую функцию, либо модифицирован, но не таким образом, как у имаго.
Кровососательный хоботок самки имаго. В первичном кровососательном ротовом аппарате самки имаго верхняя губа образует пищевой канал, мандибулы специализированы для прорезания покровов и участвуют в образовании пищевого канала, гипофаринкс от самого основания отделен от нижней губы и представляет собой стилет с каналом для проведения слюны, а максиллы участвуют в прорезании покровов (рис. 10.14.2). При этом нижняя губа (являющаяся самостоятельным лижущим хоботком) служит футляром и/или направляющим для кровососательного хоботка, оставаясь все время снаружи и не участвуя в прокалывании кожи и сосании крови (подобно нижней губе в колюще-сосущем ротовом аппарате Aphaniptera и Arthroidignatha).
Верхняя губа удлинена так, что по длине равна мандибулам и гипофаринксу, на вершине заострена и/или зазубрена (рис. 10.14.2E). На ее задней (вентральной) стороне проходит продольный желоб, служащий пищевым каналом; края этого желоба бывают сомкнуты (рис. 10.14.3B–C) или не сомкнуты; во втором случае пищевой канал замкнут благодаря прижатым к нему мандибулам (рис. 10.14.2A–B, 10.14.3D –E). При утрате мандибул функцию замыкания канала выполняет гипофаринкс (рис. 10.14.3F).
Помимо участия в кровососании, верхняя губа с пищевым каналом используется для всасывания углеводной пищи, слизываемой лабеллумами нижней губы (см. ниже). В связи с этим у большинства двукрылых, утративших кровососательную специализацию, сохраняются верхняя губа и гипофаринкс.
Каждая мандибула удлинена и уплощена, ножевидной формы с заостренной вершиной, по длине равна верхней губе и гипофаринксу (рис. 10.14.2A–С). В покое левая и правая мандибулы скрыты под верхней губой и полностью налегают друг на друга, так что режущий (исходно медиальный) край одной мандибулы совпадает с тыльным (исходно латеральным) краем другой мандибулы (рис. 10.14.2B, 10.14.3B–E). В результате этого при сокращении мышц-аддукторов (рис. 10.14.2С) режущие края мандибул выдвигаются из-под верхней губы в стороны. Благодаря режущим движениям мандибул весь кровососательный хоботок (состоящий из верхней губы, мандибул, гипофаринкса и максилл) внедряется в субстрат. Мандибулы такого строения непригодны для кусания или разгрызания пищи и способны только к прокалыванию и прорезанию покровов животных – как позвоночных, так и членистоногих. Эта специализация мандибул особенно выгодна для прокалывания толстого вязкого многослойного эпидермиса позвоночных, который, в отличие от кутикулы членистоногих, легче разрезать, чем проколоть. Одновременно с режущей функцией мандибула, лежащая дорсальнее (в разных случаях это левая или правая мандибула), служит вентральной стенкой пищевого канала. Мандибула может быть плоской, с гладкими дорсальной и вентральной поверхностями (рис. 10.14.2B), либо в средней части имеет выпуклый или вогнутый участок, комплементарный либо желобу верхней губы, либо другой мандибуле, либо дорсальной стороне гипофаринкса (рис. 10.14.3D–E). У большинства двукрылых, утративших способность к кровососанию, мандибулы имаго нефункционирующие или совсем утрачены.
Гипофаринкс сильно удлинен, склеротизован, стилетовидный, по длине равен верхней губе и мандибулам (рис. 10.14.2D, F). Слюнной канал (исходно открывающийся между гипофаринксом и нижней губой – см. Hexapoda) проходит внутри гипофаринкса (рис. 10.14.2A–B, D) и открывается на его вершине или на дорсальной стороне (рис. 10.14.3B). В кровососательном хоботке гипофаринкс расположен вентральнее сложенных мандибул (рис. 10.14.2A–C, 10.14.3B–E). В случаях утраты мандибул гипофаринкс оказывается непосредственно вентральнее верхней губы, так что его дорсальная сторона служит вентральной стенкой пищевого канала (рис. 10.14.3F), который в этом случае служит только для всасывания углеводной пищи, слизываемой лабеллумами нижней губы. В связи с этим длинный стилетовидный гипофаринкс со слюнным протоком внутри, изначально являющийся компонентом кровососательного хоботка, сохраняется у большинства двукрылых, утративших кровососательную специализацию (так же, как и модифицированная верхняя губа).
Максиллярные лацинии входят в состав колющего кровососательного хоботка, удлинены, имеют кинжаловидную или стилетовидную форму, по длине бывают равны верхней губе, мандибулам и гипофаринксу (рис. 10.14.2C), располагаются вентральнее и латеральнее гипофаринкса (рис. 10.14.2A–C, 10.14.3B –E). Исходно лацинии максилл несут зубцы и участвуют в прокалывании кожи наряду с мандибулами. У некоторых двукрылых, утративших мандибулы, таких как Asilidae и Empididae, лацинии максилл сохраняются и выполняют колюще-режущую функцию; такие двукрылые неспособны к прокалыванию кожи позвоночных и используют колющий хоботок для питания насекомыми. У многих других двукрылых, утративших кровососательную специализацию, максиллярные лацинии вестигиальные или утрачены.
Строение нижней губы имаго. Нижняя губа преобразована в лижущий хоботок, исходно приспособленный для питания углеводной пищей и одновременно служащий футляром для колюще-сосущего кровососательного хоботка (см. выше); строение нижней губы видоизменено следующим образом.
Непарная часть нижней губы на передней (дорсальной) стороне имеет глубокий желоб, в который вкладываются остальные части ротового аппарата (рис. 10.14.2F, 10.14.3A–C, F). В случае, если имеется кровососательный хоботок, он в покое скрыт в желобе нижней губы, а для кровососания освобождается из нее тем или иным способом.
Глосса и параглоссы утрачены, а основания лабиальных щупиков сближены; каждый лабиальный щупик (исходно трехчлениковый – см. Hexapoda) преобразован в нерасчлененный расширенный придаток – лабеллум (labellum).
Вентрально-медиальные стороны лабеллумов мембранозные, могут иметь псевдотрахеи. Псевдотрахея (pseudotrachea) представляет собой глубокий узкий желоб, круглый в сечении и укрепленный многочисленными поперечными склеротизованными утолщениями наподобие шпангоутов лодки; каждое такое утолщение имеет форму незамкнутого кольца, концы которого поддерживают край щели, ведущей внутрь псевдотрахеи (термин «псевдотрахея» связан с внешним сходством между этими кольцевыми утолщениями и тенидиями трахей). По направлению к основанию лабеллума псевдотрахеи впадают в общий канал (рис. 10.14.2F), так что способны под действием капиллярных сил собирать жидкость с поверхности и проводить ее к тому месту, где лабеллумы отходят от непарной части нижней губы.
У кровососущих самок во время питания углеводной пищей первичный кровососательный хоботок вложен в желоб на передней стороне нижней губы; жидкость, собираемая лабеллумами, попадает через вершину кровососательного хоботка в канал, образованный верхней губой и мандибулами (см. выше), и по этому каналу всасывается в глотку. У самцов и у некровососущих самок, утративших мандибулы, пища, собираемая лабеллумами, попадает в канал между верхней губой и гипофаринксом.
В покое лабеллумы сложены вместе так, что плоскости с псевдотрахеями обращены внутрь, друг к другу; при питании они поворачиваются этими плоскостями вентрально или в стороны.
Псевдотрахеи имеются у большинства Brachocera: у них вся мембранозная сторона каждого лабеллума исчерчена параллельными поперечными псевдотрахеями, впадающими в один продольный канал, идущий по медиальному краю (рис. 10.14.2F). Среди Nemocera такое же расположение псевдотрахей имеется у Ptychopteridae. У Tipulidae по мембранозной стороне каждого лабеллума тянутся два изогнутых канала, от каждого из которых в обе стороны ответвляется множество коротких псевдотрахей. Пара неветвящихся или двуветвистых псевдотрахей имеется у грибных комаров Mycetophilidae. У большинства других Nemocera псевдотрахеи отсутствуют.
У некоторых круглошовных мух (Cyclorapha) концы шпангоуто-подобных склеритов псевдотрахей выступают в виде склеротизованных престомальных зубцов; в этом случае нижняя губа способна не только слизывать жидкость, но также и соскребать твердые частицы и/или прорезать покровы. У этих мух первичный кровососательный хоботок утрачен, и у некоторых из них нижняя губа преобразована во вторичный кровососательный хоботок.
Зоб. От вентральной стороны стомодеума (выстланного кутикулой – см. Gnathopoda) отходит непарный дивертикул (рис. 10.14.2F). При всасывании жидкой пищи, слизываемой лабеллумами нижней губы, пища попадает в вентральный дивертикул; в дальнейшем по мере надобности эта пища перекачивается в кишечник (тогда как кровь, всасываемая при питании первичным кровососательным хоботком, сразу попадает в кишечник – см. ниже).
Гонотрофический цикл. Гонотрофический цикл выражается в четкой последовательности оплодотворения, питания кровью и созревания яиц: после оплодотворения самка активно ищет позвоночное животное, насасывает большую порцию крови, после чего происходит интенсивное развитие партии яиц; после откладки этой партии яиц самка снова готова к оплодотворению, и цикл повторяется. За свою жизнь самка может проходить один или несколько гонотрофических циклов. У многих кровососов лишь оплодотворенная самка сосет кровь; реже (у некоторых видов комаров) сосать кровь могут также неоплодотворенные самки. У многих видов комаров, мошек, мокрецов, москитов и слепней кровососание является необязательным; в этих случаях самка может отложить одну партию яиц за счет запасов белка, накопленных личинкой, но для развития следующих партий яиц ей необходимо кровососание. Таким образом, среди видов и подвидов двукрылых имеются облигатные кровососы (анаутогенные, т.е. неспособные к размножению без кровососания), факультативные кровососы (аутогенные, т.е. способные отложить одну партию яиц без кровососания) и виды, утратившие способность к кровососанию. Последние составляют подавляющее большинство видов двукрылых.
Эволюция имагинального ротового аппарата. Исходным для Diptera является глубокая специализация ротовых придатков, образующих два хоботка разного назначения (лижуще-сосущий, служащий для питания углеводной пищей, и колюще-сосущий, служащий для кровососания) и половой диморфизм, при котором кровососательный колюще-сосущий аппарат развит у самки и редуцирован у самца. Исходным для Diptera является также наличие гонотрофического цикла у самки.
Эти выводы основаны на том, что данные особенности встречаются в далеких друг от друга филогенетических ветвях двукрылых, и при этом отсутствуют у каких-либо иных насекомых. Из всех типов имагинальных ротовых аппаратов, встречающихся у двукрылых, ротовой аппарат кровососущих самок бесспорно является исходным, поскольку он включает наиболее полный набор частей первичного ротового аппарата насекомых, и только в нем имеются фунционирующие мандибулы.
Наиболее древние представители Diptera известны из триаса; жилкование крыльев на ископаемых остатках позволяет сделать вывод, что в триасе существовали представители различных современных филогенетических ветвей Diptera. Следовательно, ближайший общий предок современных двукрылых, уже имевший оба специализированных ротовых аппарата (и кровососательный, и лижущий), появился не позднее триаса. Поскольку в это время еще не существовали цветковые растения, нектар которых является важным источником углеводной пищи для современных двукрылых, триасовые двукрылые могли использовать специализированную нижнюю губу только для слизывания с растений пади – сахаристых экскрементов членистохоботных (см. Arthroidignatha). Судя по наличию лижущих лабеллумов с псевдотрахеями, нижняя губа двукрылых исходно приспособлена именно для слизывания пади, и лишь вторично используется современными двукрылыми также и для сосания нектара из цветков. Источником крови для оплодотворенных самок триасовых двукрылых могли быть существовавшие в то время разнообразные амфибии и рептилии: вероятно, триасовые виды двукрылых были также неспециализированы, как и большинство современных кровососущих видов, и питались кровью любых наземных позвоночных.
У большинства современных двукрылых первичная кровососательная специализация самок утрачена. В этих случаях источником белковой пищи для имаго может быть хищничество, детритофагия или паллинофагия; часто имаго вообще не потребляют белковую пищу. О том, что из всех видов белкового питания у имаго двукрылых кровососание является первичным, свидетельствует тот факт, что только в специализированном кровососательном ротовом аппарате сохраняются все части исходного ротового аппарата насекомых, включая действующие мандибулы, тогда как при других специализациях ротового аппарата, встречающихся у имаго двукрылых, мандибулы либо утрачены, либо не функционируют, либо сохраняют специализацию, свойственную кровососам. Ни у каких двукрылых имагинальные мандибулы не используются как грызущие. Это никак не относится к мандибулам личинок, которые у двукрылых не имеют эволюционной корреляции с мандибулами имаго и бывают как примитивными грызущими, так и различным образом модифицированными. Вероятно, исходная специализация имагинальных мандибул в качестве компонентов кровососательного аппарата столь глубока, что оказалась необратимой: в случаях изменения специализации имагинального ротового аппарата с кровососательного на какой-либо иной мандибулы неспособны изменить свою функцию, поэтому становятся бесполезными и, как правило, редуцируются.
Среди современных двукрылых первичная кровососущая специализация самок и гонотрофический цикл свойственны сравнительно небольшому числу видов, но эти виды относятся к далеким друг от друга систематическим группам. Среди Nemocera это представители по крайней мере двух неродственных таксонов – Psychodoidea и Culicoidea. Среди психодид первичными кровососами являются москиты (Phlebotomidae). Среди куликоморф первичными кровососами являются большинство видов Corethrellidae, большинство видов длиннохоботных комаров (Cilicidae), многие виды мошек (Simuliidae), часть видов мокрецов (Ceratopogonidae); предположительно также некоторые виды Chironomidae. Среди Brachocera первичные кровососы имеются только среди Tabanoidea – это отдельные виды Rhagionidae, Athericidae и большинство видов слепней (Tabanidae). У представителей этих таксонов ротовой аппарат самок имаго сохраняет действующими все части исходного ротового аппарата, включая специализированные мандибулы. У некоторых некровососущих куликоморф и табаноидов мандибулы самок также развиты. Помимо куликоморф, психодид и табаноидов, действующие мандибулы самок сохраняются у Tanyderidae и Blephariceridae; самки Blephariceridae охотятся на двукрылых и сосут их гемолимфу. У большинства двукрылых, утративших способность к кровососанию, мандибулы утратили всякую функцию: если они сохраняются, то их режущий край нежный и неспособен ничего резать (многие Simuliidae и др.). У большинства некровососущих двукрылых мандибулы совсем утрачены. Утрата мандибул происходила многократно: мандибулы утрачены у отдельных представителей названных выше таксонов, содержащих первичных кровососов, а также у многих других Nemocera и Brachocera, в том числе у всех Cyclorapha.
В таксонах, включающих кровососущих представителей, наблюдается мозаичное распределение таких признаков, как способность или неспособность самок к кровососанию и наличие или отсутствие вестигиев мандибул у самцов. При этом у самок имеются переходы между мандибулами с режущим краем, мандибулами неспособными резать и отсутствием мандибул. Если формально понимать закон о необратимости и неповторимости эволюции, то филогенетическое древо для каждого их этих таксонов и для двукрылых в целом построить невозможно. Однако следует учитывать, что в геноме общего предка двукрылых была заложена информация и о строении кровососательного аппарата с режущими мандибулами (реализуемая у самки), и об его отсутствии (реализуемая у самца). Обе эти программы были унаследованы современными двукрылыми, у которых они, как и у предков, сосуществуют в одном геноме. Поэтому в ходе эволюции самка может обратимо утрачивать режущие мандибулы (если ее ротовой аппарат строится по программе самца) и восстанавливать их (если действует программа ротового аппарата самки); самец также может обратимо приобретать мандибулы, если его ротовой аппарат уподобляется ротовому аппарату самки.
У имаго некоторых двукрылых утрачены мандибулы, но сохраняются специализированные прокалывающие максиллы; такие двукрылые не способны к кровососанию и могут использовать свой колющий хоботок только для прокалывания насекомых. Колюще-сосущий хоботок, в котором нет мандибул, а прокалывающими компонентами являются максиллы, имеется у хищных Asilidae и Empididae. У многих двукрылых, утративших первичное кровососание, не только мандибулы, но и максиллы подверглись редукции.
У большинства двукрылых нижняя губа сохраняет исходную специализацию лижуще-сосущего хоботка, который может служить как для слизывания пади с поверхности растений, так и для добывания нектара из цветков. Среди двукрылых, утративших мандибулы, многие питаются лишь углеводной пищей при помощи лижущих лабеллумов нижней губы, но некоторые вторично прибрели способность использовать лабеллумы для питания белковой пищей – пыльцой растений, насекомыми и др.
У всех круглошовных мух (Cyclorapha) первичный кровососательный хоботок утрачен, но два таксона круглошовных мух независимо друг от друга вторично перешли к питанию кровью позвоночных – это мухи-жигалки (Stomoxyinae) и живородящие мухи Pupipara Latreille 1817 {nom.typ.: Hippobosca/fg [f:1815; g:L.1758] (incl. Glossina)}. Такое вторичное кровососание принципиально отличается от первичного кровососания: в качестве прокалывающего органа здесь служит не первичный кровососательный хоботок (в котором утрачены мандибулы, но сохраняются верхняя губа и гипофаринкс), а модифицированные лабеллумы нижней губы с увеличенными престомальными зубцами. При вторичном кровососании, в отличие от первичного, отсутствует гонотрофический цикл, и кровью питаются мухи обоих полов независимо от оплодотворения.
У многих двукрылых весь имагинальный ротовой аппарат редуцирован и не функционирует.
Таким образом, взрослые двукрылые, будучи исходно нектарофагами и кровососами одновременно, могут переходить к другим типам питания и к афагии; но при этом у них не встречается настоящей фитофагии (т.е. регулярного потребление белковой пищи из живых растений). Те виды, которые имеют прокалывающий кровососательный хоботок, не используют его для прокалывания и сосания живых растений (хотя некоторые из них охотно сосут вытекающий из растений сок). В отличие от имаго, у личинок двукрылых встречается больше разных типов питания, в том числе и настоящая фитофагия.
Иные мнения об исходном строении и эволюции имагинального ротового аппарата двукрылых. Многие специалисты по двукрылым высказывают противоположное мнение о направлении эволюции имагинального ротового аппарата: по их мнению исходно имаго двукрылых были неспособны питаться кровью позвоночных. Это мнение основано на том, что подавляющее большинство современных двукрылых неспособны к кровососанию, а кровососами является сравнительно небольшая часть видов.
Однако в этом случае оказываются необъяснимыми многократное возникновение одинаковой кровососательной специализации в разных таксонах Diptera и утрата мандибул у подавляющего большинства некровососущих двукрылых.
При этом у всех остальных кровососущих насекомых, кроме двукрылых, ротовые аппараты имеют принципиально иные типы строения: у блох в прокалывании и сосании участвуют только верхняя губа и максиллярные лацинии при отсутствии мандибул (ср.: Aphaniptera); у вшей колющие стилеты образованы только нижней губой и гипофаринксом при редукции мандибул и максилл (ср.: Siphunculata); у кровососущих клопов кровососание осуществляется внутренним хоботком, состоящим из мандибул и максиллярных лациний без участия верхней губы (ср.: Arthroidignatha), а у кровососущих бабочек – хоботком, образованным галеами максилл (ср.: Glossolepidoptera). У вторичных кровососов среди двукрылых колюще-сосущий хоботок образован нижней губой. Только у кровососущих двукрылых, относящихся к парафилетическому таксону Orthorapha (см. «Классификации»), обязательными частями кровососательного хоботка являются верхняя губа и мандибулы, и только у них имеется гонотрофический цикл. Это свидетельствует о том, что все кровососущие Orthorapha унаследовали эти особенности от своего общего предка, т.е. от общего предка Diptera. Если бы эти особенности могли возникать при независимом переходе насекомых к кровососанию, они наблюдались бы не только у представителей Diptera, но и у других кровососущих насекомых.
Некоторые авторы называют некровососущих двукрылых «растительноядными» и полагают, что для двукрылых альтернативой кровососанию является «фитофагия». В действительности у двукрылых настоящая фитофагия не встречается, а питание нектаром (которое, в отличие от фитофагии, не связано с потреблением белков) происходит параллельно с кровососанием и не заменяет его.
Мнение о первичной неспособности двукрылых питаться кровью позвоночных основано лишь на предубеждении, что наличие кровососательного ротового аппарата во всех случаях является апоморфией. Такое предубеждение связано с антропоцентрическим представлением о том, что таксоны, более близкие к человеку, являются «высшими» по отношению к таксонам, более удаленным от человека, и о том, что «низшие» таксоны возникли раньше, чем «высшие». По этой логике позвоночные животные должны быть наиболее высшими, и поэтому наиболее поздними, насекомые ниже и древнее, а растения наиболее низшими и древними. Отсюда делают вывод, что для насекомых питание за счет «высших» животных должно было возникнуть позже, чем питание за счет таких «низших» организмов, как растения.
Однако в действительности дело обстоит наоборот. Из двух компонентов питания примитивного двукрылого – крови позвоночных и нектара растений – первый компонент появился на Земле гораздо раньше второго. Наземные позвоночные, пригодные для питания кровососов, существовали уже в девоне, т.е. до появления крылатых насекомых (которые известны лишь начиная с карбона). Так что для насекомых кровь позвоночных является таким же древним пищевым ресурсом, как споры растений или гифы грибов. В отличие от этого, возможность питаться нектаром растений появилась лишь в мелу, т.е. значительно позже, чем возможность питаться кровью. Поскольку двукрылые появились в триасе, они имели возможность питаться кровью позвоночных животных, но не имели возможности питаться нектаром растений.
Высказывалось предположение, что появление в мелу цветковых растений, выделяющих нектар, явилось причиной изменений в пищевой специализации двукрылых и в строении их ротового аппарата. В действительности этого быть не могло, поскольку модификации нижней губы, служащей нектарососательным хоботком, идентичны во всех филогенетических ветвях двукрылых и, следовательно, унаследованы ими от общего предка, который существовал задолго до появления цветковых растений. По всей видимости, изначально этот хоботок возник как приспособление для питания экскрементами членистохоботных, и остается таковым у современных двукрылых. Возможно, что не цветковые растения повлияли на эволюцию питания двукрылых, а наоборот, двукрылые способствовали становлению цветковых растений: будучи заранее адаптированными к питанию раствором углеводов, который они искали на растениях, двукрылые были готовы к потреблению нектара еще до его появления. В этой ситуации выделение растением нектара вблизи генеративных органов сразу приводило к их опылению, что способствовало эволюции энтомофильных цветков.
● Модификация крылового аппарата. Крыловой аппарат переднемоторный: для полета служат только передние крылья, тогда как задние крылья превращены в жужжальца (см. ниже). В связи с этим среднегрудь (вмещающая мышцы, двигающие передние крылья) сильно увеличена, с крупным скутумом, а переднегрудь и заднегрудь уменьшены и соединены с ней неподвижно.
Переднее крыло в проксимальной части сужено в виде б.м. выраженного стебелька с модифицированным жилкованием (рис. 10.14.1A–B).
Задние крылья превращены в жужжальца (halteres) – придатки гантелевидной формы: жужжальце подвижно причленено к туловищу узким основанием, в проксимальной части утолщено, в средней части тонкое и цилиндрическое, на вершине утолщено (рис. 10.14.1B). Утолщенная проксимальная часть жужжальца несет хордотональные сенсорные органы. В полете жужжальца совершают самостоятельные быстрые вибрирующие движения. В отличие от многих переднемоторных насекомых, у Diptera жужжальца никогда не сцепляются с передними крыльями.
Жужжальца выполняют роль гироскопа, помогающего летящему насекомому ориентироваться в пространстве. Благодаря этому определенные представители двукрылых, в частности многие Cyclorapha, способны к очень быстрому и маневренному полету. Однако такие же жужжальца имеются у всех двукрылых, включая примитивных Nemocera, полет которых медленный и менее маневренный, чем у многих других насекомых. Поэтому исходное значение жужжалец не вполне ясно.
У куколки задний протоптерон (придаток, онтогенетически гомологичный жужжальцу имаго) расположен так же, как задний протоптерон других насекомых – направлен вентрально и в значительной мере закрыт передним протоптероном [см. Metabola (1Ba)] (рис. 10.14.5B, F).
Обычно крылья развиты у обоих полов, реже уменьшены или утрачены; иногда крылья развиты у самца и уменьшены или утрачены у самки. Жужжальца сохраняются у всех Diptera, имеющих развитые передние крылья, и у части представителей, утративших передние крылья; лишь у некоторых Diptera, полностью утративших крылья, жужжальца также утрачены.
Наличие жужжалец является уникальной аутапоморфией Diptera, не встречающейся ни у каких других насекомых. Иногда «жужжальцами» называют уменьшенные задние крылья самцов кокцид (Gallinsecta) и некоторых других насекомых. В отличие от настоящих жужжалец двукрылых, уменьшенные задние крылья этих насекомых либо сцепляются с передними (рис. 9.12.1E), либо неспособны к активным движениям, либо то и другое.
● Личинки безногие. Личиночные ноги [не имеющие эволюционной корреляции с имагинальными – см. Metabola (1Ccα)] редуцированы так, что на месте ноги ничего не выступает, и внешне личинка кажется совершенно безногой. Однако сохраняются вестигии ног: каждая личиночная нога представлена только своей вершиной, которая образует участок стенки груди и несет группу из 4 щетинок, расположенных как бы по углам квадрата (рис. 10.14.4A–B). Все это образование, гомологичное личиночной ноге других Metabola, обозначают как «d-орган» (Keilin 1915), или орган Кейлина. Иногда его называют внутренним зачатком куколочной ноги; однако в состав d-органа входит участок наружной кутикулы, соответствующий ему участок гиподермы, четыре щетинки и соответствующие им сенсорные и оболочечные клетки – то есть этот орган является не только внутренним, но и наружным, и не сводится к куколочному зачатку, а содержит также сугубо личиночные детали строения (щетинки), отсутствующие у куколки. Четыре щетинки d-органа бывают либо раздельными (рис. 10.14.4A–B, E–G), либо слиты вместе, сохраняя четыре основания (рис. 10.14.4D), либо модифицированы иным образом; иногда щетинок меньше четырех, иногда все щетинки утрачены. Кутикула d-органа либо неотличима от кутикулы туловища, либо отличается по толщине и рельефу.
Гиподерма d-органа может вдаваться внутрь тела (рис. 10.14.4D). У комаров-долгоножек (Tipulidae) карманы d-органов глубокие и стебельчатые, так что гиподерма вестигия ноги полностью погружена в карман, а ее вершина не достигает покрова (но от нее идут четыре сенсорные реснички к четырем щетинкам на поверхности тела) (рис. 10.14.4F–G). У круглошовных мух (Cyclorapha) d-органы также имеют глубокие стебельчатые карманы и известны как ножные «имагинальные диски».
При превращении личинки в куколку куколочные ноги всегда образуются из d-органов [т.к. куколочная нога всегда развивается из личиночной ноги или ее вестигия – см. Metabola (1Cc)].
● Модификация дыхалец. На всех стадиях развития сохраняются все 10 пар трахеостий (см. Amyocerata), но при этом число действующих дыхалец различно у личинки, куколки и имаго.
У личинки трахеостии последней, VIII абдоминальной, пары (называемые постабдоминальными) отличаются от остальных и обычно представляют собой действующие дыхальца; иногда это единственная пара дыхалец; если личинка имеет и другие действующие дыхальца, то дыхальца VIII абдоминальной пары первыми возникают в онтогенезе и обычно являются самыми крупными.
Куколка исходно имеет лишь одну пару действующих дыхалец – стеноторакальную (иногда неверно называемую «переднегрудной»); каждое дыхальце открывается на вершине особой дыхательной трубки (рис. 10.14.5A, C, E). У куколок некоторых двукрылых, помимо стеноторакальных, имеются те или иные пары абдоминальных дыхалец (рис. 10.14.5A). Куколки, обитающие под водой, используют трубки для дыхания воздухом (рис. 10.14.5C, E), но такие же трубки имеются и у куколок, обитающих вне воды (рис. 10.14.5A).
У имаго самцов VIII абдоминальные трахеостии всегда замкнутые и нефункционирующие (хотя у личинки именно они обычно являются единственными дыхальцами). У имаго бывают замкнуты и некоторые другие пары трахеостий.
Вероятно, исходно личинки двукрылых адаптированы к полуводному образу жизни, при котором бóльшую часть времени задний конец тела, несущий единственную пару дыхалец, сообщается с воздушной средой, а остальное тело, лишенное дыхалец, погружено в жидкость. Личинки многих двукрылых сохранили такой образ жизни, обитая на поверхности и/или у берега водоемов, во влажных наземных биотопах, в органических остатках или в тканях живых растений или животных. Во многих случаях личинки перешли к обитанию в других средах – наземной или, наоборот, водной; при этом их дыхательная система либо сохраняет исходное строение, либо изменена, адаптировавшись к новым условиям. Судя по особому расположению дыхалец у куколок, куколки двукрылых также исходно адаптированы к обитанию на границе воды и воздуха; при переходе к другим средам обитания дыхательная система куколок либо сохраняет исходное строение, либо изменена.
● Склеенная куколка. Куколка исходно склеенного типа [pupa conglutinata – см. Metabola (1Bb)] (рис. 10.14.5); среди двукрылых лишь у Cyclorapha куколка вторично несклеенная (pupa adectica exarata), что связано с наличием пупария (см. ниже)].
Признаки Diptera неясного филогенетического значения
(7) Число антенномеров. У имаго флагеллум антенны предположительно исходно 14-члениковый [т.е. антенна исходно 16-члениковая – см. Amyocerata (1)]. У различных Nemocera число флагелломеров бывает от 3 до 14. У Brachocera число флагелломеров сокращено до 8; бывает еще более сокращено или вторично увеличено [см. ниже, Brachocera (1)].
Классификации Diptera
I. Деление Diptera на Orthorapha и Cyclorapha. Таксон Orthorapha Brauer 1863 – прямошовные (другие написания: Orthorhapha Brauer 1880 = Orthorrhapha Sharp 1894) представляет собой заведомо парафилетический таксон и характеризуется лишь отсутствием аутапоморфий, свойственных Cyclorapha. Название «Orthorapha» связано с тем, что, в отличие от Cyclorapha, у Orthorapha куколка не заключена в пупарий, открывающийся по кольцевому шву; у большинства Orthorapha куколка ничем не закрыта, и поэтому при вылуплении имаго видно, что куколочная кутикула трескается по прямому продольному шву на дорсальной стороне, как и при большинстве линек у насекомых (см. Hexapoda).
Таксон Cyclorapha является голофилетическим (см. ниже).
Данная классификация оправдана только тем, что голофилетический таксон Cyclorapha по числу видов значительно превосходит парафилетический таксон Orthorapha; однако она во всех отношениях менее оправдана, чем предложенное ранее деление Diptera на Nemocera и Brachocera.
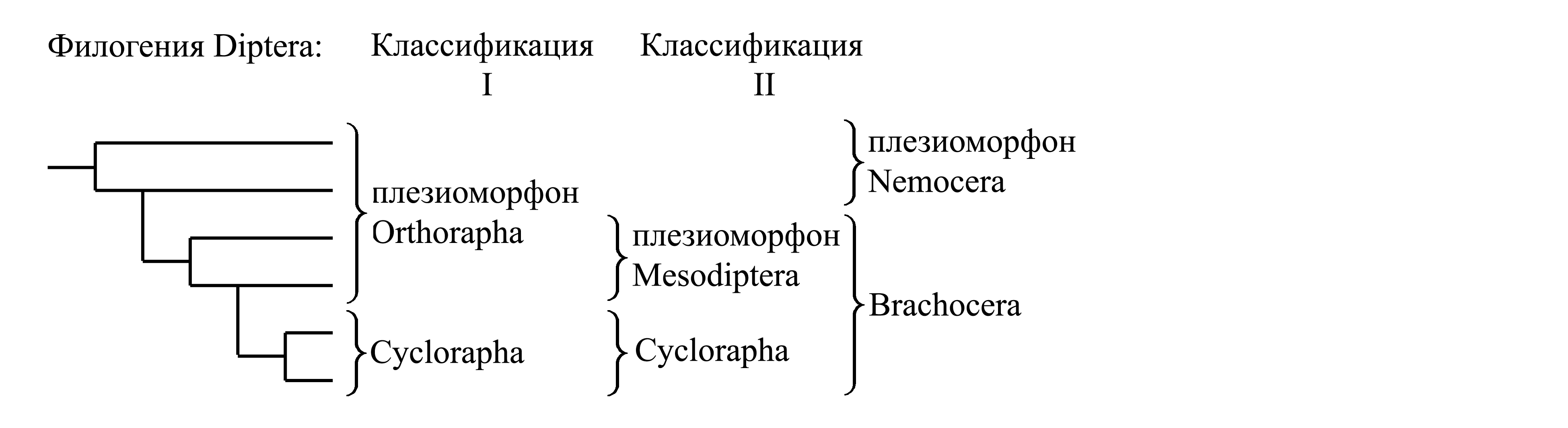
II. Деление Diptera на Nemocera и Brachocera. Таксон Nemocera представляет собой плезиоморфон, меньший по числу видов, чем голофилетический таксон Brachocera. Существует несколько попыток расформировать плезиоморфоны Nemocera и Mesodiptera, однако ни одна из них не стала общепризнанной.
|
1. Плезиоморфон Nemocera Latreille 1817 NomCirc. – длинноусые, или комары в широком смысле (syn.circ.: Polyarta Billberg 1820 = Nematocera Berthold 1827 (non Nematocera Dumeril 1805, nec Nematocera Meigen 1818) = Polymera Burmeister 1829 = Nemocerae Gray 1832 = Nemocerata Perty 1833 = Nematocerata Gravenhorst 1843 = Macroceratae Bigot 1885 = Nemoceratae Bigot 1888 = Nematoceratae Bigot 1892 = Prodiptera Crampton 1916a = Nematocerina Pearce 1936; nom.typ.: Tipula/fg [f:1802; g:L.1758(gen.221)] (incl. Culex, Bibio)}. У имаго флагеллум антенны содержит от 3 до 14 члеников; строение антенн разнообразно, но, в отличие от Brachocera, флагеллум не подразделен на постпедицеллус и стилус. Максиллярные щупики имаго бывают 5-члениковыми или 4-члениковыми. Ротовой аппарат личинок либо грызущий, либо так или иначе модифицирован, но не такой, как у Brachocera. Длина тела 0,5–60 мм. Около 50 000 видов; всесветно. WWWWWWWWWWWWWWWWWWWWWW |
|
2. Brachocera Macquart 1834 NomCirc. – короткоусые, или мухи в широком смысле {syn.circ.: Brachycera Zetterstedt 1842 = Brachycerina Pearce 1936; nom.hier.: Musca/f2=Oestrus/g2 [f:1781; g:L.1758(gen.220)] (sine Tipula; incl. Tabanus)}. Характеризуется следующими аутапоморфиями.
● У имаго антенны уменьшены, а число флагелломеров сокращено до 8; проксимальная часть флагеллума (называемая постпедицеллус – postpedicellus) дифференцирована как хеморецепторная: флагелломеры в той или иной мере утолщены и слиты; дистальная часть флагеллума (называемая стилус – stylus) дифференцирована как механорецепторная: флагелломеры в той или иной мере утоньшены (рис. 10.14.1B) (тогда как в исходной антенне механорецепторным является сочленение флагеллума с педицеллусом – см. Amyocerata). У различных Brachocera постпедицеллус может быть 1-члениковым, а стилус 7-члениковым; иногда постпедицеллус состоит из большего числа члеников, а стилус из меньшего (Stuckenberg 1999). В частности, у Cyclorapha постпедицеллус особенно крупный, слитный и неподвижный, а стилус, называемый ариста (arista), 3-члениковый.
● У имаго максиллярные щупики (исходно пятичлениковые – см. Hexapoda) направлены вперед и функционально одночлениковые: каждый состоит из 2 неподвижно соединенных члеников или из одного членика. Двучлениковое строение максиллярных щупиков сохраняется у некоторых Mesodiptera (рис. 10.14.2C); у всех Cyclorapha максиллярные щупики одночлениковые.
● У личинки ротовой аппарат преобразован (рис. 10.14.4A, C) следующим образом. Основания мандибул смещены дорсально, так что мандибулы направлены остриями вентрально и двигаются не навстречу друг другу, а параллельно друг другу в дорсо-вентральном направлении. Верхняя губа (исходно уплощенная дорсовентрально), будучи расположенной между сближенными мандибулами, приобрела сжатую с боков форму (у Mesodiptera верхняя губа может быть хорошо развита, у Cyclorapha сохраняется только в первом возрасте). Максиллярные лацинии направлены, как и мандибулы, дорсовентрально, в той или иной мере прирастают к мандибулам, десклеротизованы, иногда (в частности у Cyclorapha) совсем слиты с мандибулами и не выражены. Расположенные латеральнее каждой мандибулы максиллярный щупик и антенна сближены между собой.
Головная капсула в той или иной мере погружена в туловище: иногда в туловище погружена лишь задняя часть головы, иногда вся голова способна втягиваться в туловище (рис. 10.14.4A, C).
2.1. Плезиоморфон Mesodiptera Crampton 1916a NomCirc. {syn.circ.: Orthobrachycera Brunetti 1912; по циркумскрипции также соответствует: «Brachycera» sensu Brauer 1880 (non Zetterstedt 1842); «Orthorrhapha» auct. (non Sharp 1894; nec Orthorapha Brauer 1863); « Brachycera-Orthorrhapha» auct.; nom.typ.: Bombylius/f1=Tabanus/g1 [f:1802:427; g:L.1758(gen.223)]}. В отличие от Cyclorapha, всегда сохраняется головная капсула личинки (частично или полностью втянутая в грудь – см. Brachocera); имагинальные диски отсутствуют; имеется не менее четырех личиночных возрастов; при линьке личинки на куколку личиночная кутикула обычно сбрасывается, так что куколка не заключена пупарий, склеенная. От нескольких миллиметров до 8 см (некоторые Asilidae). Около 37 000 видов (Zhang 2011); всесветно. WWWWWWWWWWWWWWWWWWWWWW |
Kluge0279.JPG) |
2.2. Cyclorapha Brauer 1863 NomCirc. – круглошовные {syn.circ.: Cyclorhapha Schiner 1864 = Discota Weismann 1866 = Cyclorrhapha Becher 1882; nom.hier.: Musca/f3=Oestrus/g3 [f:1781; g:L.1758(gen.220)] (sine Tabanus; incl. Lonchoptera). Характеризуется следующими аутапоморфиями: у личинки (безногой – см. Diptera) головная капсула утрачена, а ротовой аппарат сильно модифицирован (см. ниже); при превращении личинки в куколку образующиеся в ее голове и груди предлиночные складки гиподермы имеют форму своеобразных «имагинальных дисков» (см. ниже); процесс превращения личинки в куколку сопровождается образованием пупария (см. ниже).
Личинка имеет лишь 3 возраста (в отличие от 4–10 возрастов у других двукрылых).
Головная капсула личинки (исходно втянутая в грудь – см. выше, Brachocera) полностью утрачена. Внешняя часть головы маленькая, мембранозная, несет лишь пару антенномаксиллярных выступов (см. выше, Brachocera), на которых сидят очень маленькие мембранозные вестигии антенн (дорсальные папиллы) и максиллярных щупиков (вентральные папиллы). Мандибулы (двигающиеся дорсовентрально – см. Brachocera), гипофаринкс и тенториум преобразованы в уникальный ротоглоточный аппарат: задние руки тенториума утрачены, а передние полностью слиты с модифицированными глоточными склеритами, образуя внутренний скелет, к которому спереди причленяется пара ротовых крючьев – мандибул. При выдвижении ротоглоточного аппарата вперед мандибулы выдвигаются наружу, а при отведении ротоглоточного аппарата назад мандибулы полностью погружаются в туловище.
У личинки третьего (последнего) возраста внутри тела имеются несколько парных и непарных округлых телец, названных «имагинальными дисками» (эпитет «имагинальный» здесь неверен, поскольку из этих дисков развиваются не имагинальные, а куколочные придатки). Каждый «имагинальный диск» образован ввернутым участком гиподермы с вдающимся в него зачатком куколочного придатка; по своему строению имагинальный диск Cyclorapha похож на зачаток куколочной ноги у комара-долгоножки (рис. 10.14.4F–G), но отличается тем, что может утрачивать связь с внешней гиподермой. Имеются три пары дисков ног, пара дисков стеноторакальных дыхательных трубок (см. выше), две пары дисков протоптеронов, а также диски головы, антенн и хоботка. Диски могут срастаться с трахеями (которые также образованы эктодермальным эпителием) и друг с другом, а также образовывать вторичное срастание с наружным эпителием, так что бывает трудно найти место, от которого отделился диск. При превращении личинки в куколку (происходящем без сбрасывания кутикулы – см. ниже) устье диска раскрывается, содержащийся в нем придаток оказывается наружными, а эпителий диска, окружавший придаток, входит в состав наружной стенки туловища.
По своему происхождению, развитию и функциям диски принципиально различны.
Три пары дисков ног представляют собой модифицированные личиночные сенсорные d-органы (см. выше); как и d-органы других двукрылых, они существуют на протяжении всего личиночного развития, а при метаморфозе превращаются в куколочные ноги. В отличие от многих других двукрылых, у Cyclorapha d-орган несет не четыре, а лишь три маленькие щетинки. Соответственно, клетки d-органа участвуют в образовании покровных кутикулярных структур – по крайней мере, образуют эти три щетинки. После формирования кутикулы и щетинок третьего (последнего) личиночного возраста гиподерма d-органа вворачивается, образуя стенку замкнутого диска, и погружается в туловище; нервные клетки d-органа утрачивают связь с принадлежащими им тремя щетинками. Таким образом, ножные диски представляют собой модифицированные предлиночные складки, образующиеся при превращении личиночных ног (d-органов) в куколочные.
Передняя пара дорсальных дисков появляется у личинки только в последнем возрасте, когда гиподерма начинает формировать куколочные зачатки. Это предлиночные складки, из которых развиваются куколочные стеноторакальные дыхательные трубки.
Вторая и третья пары дорсальных дисков – диски протоптеронов – закладываются раньше третьего возраста и переходят из возраста в возраст, оставаясь внутренними образованиями. Подобные внутренние зачатки протоптеронов, вероятно, независимо возникли у разных насекомых с полным превращением [см. Metabola (1Cd)].
Имагинальные диски головы и ее придатков оказались внутренними образованиями в силу того, что у личинки Cyclorapha ткани, соответствующие тканям головы, полостью погружены в туловище; соответственно и развивающиеся из них ткани куколочной головы оказываются внутри туловища личинки.
Термин «имагинальные диски» (по-немецки «Imaginascheiben») был введен Вейсманном (Weismann 1864) в том значении, которое приведено выше. В связи с этим было предложено название Discota Weismann 1866 для группы насекомых, включающей мух (Cyclorapha) и противопоставляемой Adiscota Weismann 1866, куда были включены комары (Culicidae).
Позже некоторые авторы стали называть «имагинальными дисками» не только внутренние замкнутые гиподермальные образования, но и любые участки гиподермы (или даже любой ткани), которые принимают участие в образовании каких-либо имагинальных тканей, и не только у мух, а и у других насекомых, включая комаров. В таком случае считают, что у личинок мух имагинальные диски имеются не только в груди, но и в брюшке. При этом под «имагинальными дисками» брюшка подразумевают участки обычной наружной личиночной гиподермы (образующей нормальную личиночную, а затем и куколочную кутикулу), которые при трансформации куколки в имаго разрастаются и образуют имагинальную гиподерму, в то время как другие участки личиночно-куколочной гиподермы вытесняются растущими «имагинальными дисками» и лизируются. С тем же основанием можно применительно к любому многоклеточному животному называть «имагинальными дисками» любые участки тканей, клетки которых не дегенерируют в ходе онтогенеза. Путаное употребление термина «имагинальные диски» приводит к недоразумениям, поскольку, с одной стороны, считают наличие «имагинальных дисков» особенностью личинок Cyclorapha (что соответствует только исходному значению этого термина), а с другой стороны, тех же у личинок Cyclorapha под этим же термином описывают то, что имеется у любого животного.
У закончившей развитие личинки III (последнего) возраста покровы не сбрасываются, а меняют форму, склеротизуются и образуют пупарий [вместилище для куколки – см. Metabola (1D)]. Куколка находится внутри пупария. Дыхательные трубки куколки (стеноторакальные – см. выше) обычно срастаются со стенкой пупария и проходят сквозь нее в том месте пупария, которое образовалось из кутикулы первого сегмента брюшка личинки; в некоторых таксонах дыхательные трубки не соединяются со стенкой пупария и их дыхательные отверстия открываются внутри пупария. Когда взрослая муха, вылупившаяся из куколки, выходит из пупария, покров пупария трескается по определенным линиям. У большинства Cyclorapha эти линии образуют замкнутый контур, так что от пупария отваливается часть покрова в виде крышечки. Название «Cyclorapha» («круглошовные») связано с замкнутой формой шва, по которому открывается пупарий. Исключение представляют Lonchopteridae {nom.circ. Oxypterae Meigen 1824 = Acroptera Brauer 1869 = Anatria Meijere 1900; nom.typ.: Lonchoptera/fg [f:1835; g:1803]}, у представителей которого пупарий открывается незамкнутым швом Т-образной формы, так что крышечка не отваливается.
Длина тела 0,5–25 мм. Около 63 000 видов (Zhang 2011); всесветно.
* * *
KlugeIMGP0901.JPG) |
Kluge0262.JPG) |
Подробнее см. Н.Ю. Клюге 2020. «Систематика насекомых и принципы кладоэндезиса».
Kluge0192.JPG)