КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ

![]()
|
КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VIII-1.2.2.4. Dermatoptera Burmeister 1838 – уховертки
(Somobialia
Eucaryota Metazoa Monostomata
Haemataria Peritoneata
Metameria
Gnathopoda Euarthropoda
Mandibulata Atelocerata
Hexapoda Amyocerata
Pterygota
Metapterygota Neoptera
Rhipineoptera Tegminoptera
Dermatoptera)
(рис. 8.7.1–8.7.3)
(рис. 8.7.1,
8.7.2, 8.7.3)
|
Названия, соответствующие по циркумскрипции: — Labidoura Dumeril 1805 (исходно только во французском написании Labidoures и в немецком написании Labidouren); — Placoda Billberg 1820; — Trimera Zetterstedt 1821 (non Trimera Dumeril 1805); — «Euplekoptera» Westwood 1831 (с опечаткой); — Euplexoptera Westwood 1838; — Dermatoptera Burmeister 1838; NomCirc. — Labidura (Burmeister 1838) Haeckel 1866 (non genus Labidura Leach 1815); — Dermoptera Agassiz 1848; — Harmoptera Fieber 1852; — Euplectoptera Fischer 1853 (non Euplectoptera Tillyard 1932); — Pseudoorthoptera Dominique 1893 (исходно только во французском написании Pseudoorthoptères); — Dermaptoria Blatchley 1920. По циркумскрипции также соответствует: — «Dermaptera» sensu Kirby 1813 (non De Geer 1773); — таксонам под родовым названием Forficula (sensu Linnaeus 1758) и производными типифицированными названиями: Forficulariae, Forficulaedes, Forficulidae, Forficularia, Forficulites, Forficulina, Forficulodea, Forficulida, Forficulidi. Иерархическое название: Forficula/f1=g3 [f:1810; g:L.1758(gen.192)]. |
|
Ранг. Отряд или надотряд.
Названия. Для этого таксона часто употребляют название «Dermaptera» с авторством «De Geer 1773». В действительности исходно таксон Dermaptera De Geer 1773 {syn.circ.: Ulonata Fabricius 1775} включал роды Mantis, Locusta, Acrydium, Gryllus, Blatta и Forficula (что соответствует нынешним таксонам Raptoriae, Spectra, Saltatoria, Neoblattariae и Dermatoptera) (см. Tegminoptera: «Название»). Позже Ретзиус (Retzius 1783), использовавший классификацию Де Геера, переименовал Dermaptera в Hemiptera (о разных значениях названия Hemiptera см. ниже, Arthroidignatha: «Название»), а название Dermaptera употребил для таксона, ныне называемого Heteroptera (см. ниже). Kirby (1813) предложил употреблять название Dermaptera применительно только к уховерткам (что соответствует только роду Forficula в классификации Де Геера). Для того, чтобы соблюсти циркумскриптный принцип номенклатуры, Westwood (1831) предложил для уховерток новое название Euplexoptera. Другим названием, соответствующим по циркумскрипции, является Dermatoptera Burmeister 1838; несмотря на то, что это название младше, оно мне кажется более удобным, т.к. сходно с широко известным, но неверным названием «Dermaptera».
Характеристика Dermatoptera (рис. 8.7.1)
Голова прогнатная; в сочленении головы с туловищем, помимо пары латеральных шейных склеритов (см. Pterygota2), имеются два дополнительных вентральных шейных склерита (рис. 8.7.1A). Простые глазки всегда отсутствуют (см. Amyocerata). Глоссы нижней губы (см. Hexapoda) утрачены, так что имеется лишь одна пара апикальных придатков – параглоссы; в остальном ротовой аппарат обычного грызущего строения (см. Rhipineoptera).
Модификации крылового аппарата. Крыловой аппарат (исходно заднемоторный, с уплотненными передними крыльями и анальным веером на задних крыльях – см. Tegminoptera) сильно видоизменен: задние крылья имеют уникальную модификацию анального веера, позволяющую складывать его не только веером, но и поперек, а передние крылья укорочены и приобрели аппарат, позволяющий фиксировать их на спине.
Передние крылья (имеющие тенденцию к элитризации – см. Tegminoptera) превращены в плотные надкрылья (тегмины) (рис. 8.7.1A). Анальный край надкрылья прямой, и в покое надкрылья сложены на спине так, что их анальные края сходятся и налегают друг на друга лишь узкими полосками. Сложенные надкрылья фиксируются на спине благодаря особому приспособлению: на вентральной стороне каждого надкрылья вдоль его анального края тянется продольный гребень, несущий крепкие щетинки, направленные латерально-назад; на скутуме заднегруди имеется продольное медиальное вдавление, обрамленное парой продольных гребней, несущих крепкие щетинки, направленные медиально-назад (рис. 8.7.1C). У вымерших Protelytroptera надкрылья закрывают все брюшко; у Brachydermaptera (куда входят все современные уховертки) надкрылья сильно укорочены; жилкование надкрыльев сохраняется у вымерших Protelytroptera и Archidermaptera (рис. 8.7.2B, C, E) и утрачено у всех современных уховерток.
Заднее крыло имеет следующее строение (рис. 8.7.1). Непереворачивающаяся бласть крыла впереди анального веера преобразована в так называемый фустис (fustis): он сильно уменьшен, склеротизован, с обедненным жилкованием. У полнокрылых форм при сложенных крыльях дистальная часть фустиса всегда выступает сзади из-под короткого надкрылья (рис. 8.7.2C). Инверталярная область длиннее фустиса, так что вершинная часть крыла несет только веерообразно расположенные ваннальные жилки; эта вершинная часть крыла с ваннальными жилками способна складываться, сгибаясь вперед. Проксимальные части ваннальных жилок слиты друг с другом таким образом, что анальный веер расходится не от основания крыла, а имеет вид либо гребенки (у вымерших Protelytroptera) (рис. 8.7.2F), либо веера, расходящегося из точки, сильно удаленной от основания крыла (у Brachydermaptera) (рис. 8.7.1A). У всех современных уховерток от вершины фустиса веером расходятся 8 ваннальных жилок (V1 –V8); передние из них в расправленном виде направлены дистально, так что крыло с расправленными ваннальными жилками значительно длиннее фустиса. Позади них от общего ваннального ствола отходят порознь еще две ваннальные жилки (V9 –V10). Между каждыми двумя соседними ваннальными жилками находится интеркалярная жилка, имеющая свободное основание. Все ваннальные и интеркалярные жилки соединены одной цепью поперечных жилок, каждая из которых имеет слабый участок, по которому при складывании веера проходит радиальная складка. Проксимальнее цепи поперечных жилок каждая ваннальная и интеркалярная жилка имеет уплощенный и расширенный участок характерной изогнутой формы; у ваннальных и интеркалярных жилок изгибы уплощенных участков направлены в противоположные стороны. При складывании крыла анальный веер складывается так, что все интеркалярные жилки переворачиваются; сложенный веер сгибается пополам по линии, проходящей по уплощенным расширенным участкам жилок; это происходит в результате того, что по мере отведения жилок назад их дистальные участки отгибаются вперед. Таким образом сложенное крыло оказывается по длине равно его фустису и может быть в три раза короче расправленного крыла и в два раза короче каждой ваннальной жилки.
У личинок протоптероны выражены только в последних возрастах и имеют характерное строение (рис. 8.7.2A).
Особенности ног. Шпоры на вершине голени утрачены (см. Neoptera). Лапки (исходно пятичлениковые – см. Amyocerata) у всех современных уховерток 3-члениковые (рис. 8.7.1A), но у примитивных вымерших Archidermaptera лапки достоверно сохраняли все 5 члеников (рис. 8.7.2B–C).
Особенности брюшка. Благодаря тому, что передние крылья укорочены, а задние крылья компактно складываются (см. выше), сложенные крылья не закрывают брюшко. Сегменты брюшка соединены подвижно, и брюшко способно сильно изгибаться; при этом сочленовные мембраны брюшка защищены налегающими краями тергитов и стернитов. Задние края тергитов и стернитов брюшка широко налегают на позади лежащие тергиты и стерниты, а боковые края тергитов (параноты) налегают на стерниты; исключение составляют VIII–IX уромеры самки (см. ниже) (рис. 8.7.1A). В результате этого каждая пара абдоминальных дыхалец (находящихся на плеврах I–VIII уромеров – см. Amyocerata) закрыта паранотами своего сегмента, а поверх них – еще налегающими задними краями впереди лежащего стернита, так что снаружи дыхальца не видны. У самца видны все 10 тергитов брюшка; у самки видны только 8 тергитов брюшка (I–VII и X) (рис. 8.7.1A, 8.7.3E); внешне вершина брюшка кажется похожей у самца и самки, поскольку у обоих полов десятый сегмент увеличен и с вентральной стороны прикрыт удлиненным стернитом предыдущего видимого сегмента (который является девятым у самца и седьмым у самки).
Брюшко самки. У самки VIII и IX уромеры (исходно несущие яйцеклад – см. Amyocerata) сильно укорочены и неподвижно слиты с десятым уромером (рис. 8.7.3F). Тергит и стернит VII уромера полностью закрывают VIII и IX уромеры; при этом стернит VII уромера увеличен, простирается назад и с вентральной стороны налегает на X уромер (рис. 8.7.1A, 8.7.3E). Так что внешне брюшко самки кажется состоящим из 8 тергитов (II–VII и X) и 6 стернитов (II–VII) (поскольку I стернит редуцирова)
В отличие от имаго, у молодых личинок самок VIII и IX уромеры вполне развиты, подвижны и не скрыты, так что все уромеры равны по длине, как и у самцов.
В результате того, что у взрослых самок VIII и IX уромеры укорочены и скрыты, первичный яйцеклад не может отгибаться вентрально и сохранять свои исходные функции. У вымерших мезозойских Archidermaptera яйцеклад выглядит как вполне развитый: длинный, прямой, состоящий из сомкнутых створок (рис. 8.7.2B). У всех современных уховерток яйцеклад утратил олистетеры (рис. 8.7.3E–F), а у большинства уховерток совсем утрачен (рис. 8.7.1A).
Копулятивный аппарат самца. Гениталии самца (расположенные на границе XI и X уромеров – см. Amyocerata) имеют сложное строение (рис. 8.7.3B–D), способны втягиваться в брюшко и выдвигаться из него; у многих уховерток семяизвергательный канал разделен на пару каналов (рис. 8.7.3С), а у некоторых каждый из них разветвлен еще на два, т.е. имеется до 4 семяизвергаткльных отверстий.
Модификация церков. У современных уховерток церки (только парные – см. Metapterygota) одночлениковые и превращены в хватательные придатки клещевидной формы. Мышцы, идущие от оснований церков к X тергиту брюшка (см. Amyocerata), направлены не только вперед, но и назад: от основания каждого церка назад отходит мышца-раздвигатель, которая крепится в особом заднем выступе тергита, простирающимся между церками; у большинства взрослых уховерток этот выступ, называемый пигидий (pygidium), подвижно сочленен с остальной частью тергита (рис. 8.7.3B).
У подавляющего большинства современных уховерток церки имаго склеротизованные, хватательные, с заостренными вершинами, изогнутыми медиально (рис. 8.7.1A, 8.7.3E), часто с зубцами на медиальной стороне. Уховертки могут использовать клещевидные церки для ловли добычи, что возможно благодаря длинному гибкому брюшку, не закрытому надкрыльями и способному загибаться вверх и вперед до головы. Форма церков видоспецифична и часто различается у самца и самки; церки самца обычно крупнее и имеют более сложную форму.
У личинок церки менее специализированы, чем у имаго. У наиболее примитивных уховерток церки личинки не клещевидные, а прямые, длинные, многочлениковые, с неопределенно большим числом члеников (рис. 8.7.3A). При этом у зрелой личинки самый проксимальный членик значительно длиннее других, и при линьку на имаго из него развивается одночлениковый клещевидный церк имаго; остальную часть церка личинка перед линькой на имаго может съедать (Green 1898, Simizu & Machida 2011). Церки личинки обычно способны к автотомии: около самого основания церк имеет слабое место, по которому он легко отламывается (рис. 8.7.3A); у имаго, наоборот, церки очень прочные.
У вымерших палеозойских Archidermaptera церки имаго были многочлениковыми, либо прямыми (рис. 8.7.2B–С), либо с клещевидным первым члеником (рис. 8.7.2D),.
Классификация Dermatoptera с учетом вымерших групп
Филогения Dermatoptera представляется следующим образом:
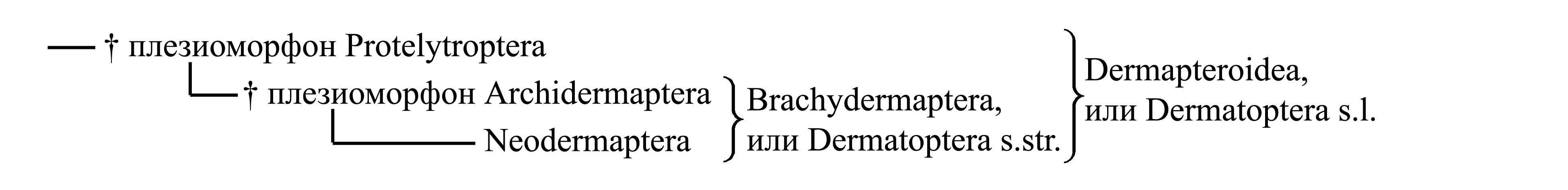
1. Dermapteroidea Jeannel 1949a NomCirc. {syn.circ.: Dermapteroida Kevan & Knipper 1961}, или Dermatoptera в самом широком смысле. Помимо настоящих уховерток, включает вымерших Protelytroptera.
1.1. † Плезиоморфон Protelytroptera Tillyard 1931a NomCirc. {syn.circ.: Protocoleoptera Tillyard 1924b = Elytroptera Tillyard 1931a (non Elytroptera Clairville 1798); nom.typ. Protelytron/fg [fg:1931]}. По внешнему виду напоминают жуков: тело не удлинено, овальное; надкрылья не укорочены, закрывают все брюшко и сложенные задние крылья. Заднее крыло значительно длиннее и шире переднего (рис. 8.7.2E–F); в покое складывается веерообразно, а его вершинная часть сгибается, так что заднее крыло полностью скрывается под надкрыльем. Лапки 5-члениковые. Церки короткие, членистые. Вымершая группа, существовавшая в перми (рис. 2.2).
1.2. Brachydermaptera Kluge 2003 NomCirc., или Dermatoptera в более узком смысле. Передние крылья сильно укорочены, а задние складываются особым образом так, что бóльшая часть брюшка остается не закрытой крыльями (см. выше). Помимо современных уховерток, включает вымерших примитивных вымерших Archidermaptera.
1.2.1. † Плезиоморфон Archidermaptera Bey-Bienko 1936 NomCirc. {nom.typ.: Protodiplatys/fg [fg:1925]} (рис. 8.7.2B–C). Лапки 5-члениковые. На надкрыльях могут быть ясно выраженные продольные жилки (рис. 8.7.2B–C). Церки взрослых насекомых многочлениковые, а десятый сегмент брюшка не увеличен. Вымершая группа, существовавшая в мезозое (рис. 2.2).
1.2.2. Neodermaptera Engel 2003 NomCirc., или Dermatoptera в еще более узком смысле. Лапки 3-члениковые. На надкрыльях жилкование полностью утрачено. Церки по крайней мере у имаго нечленистые. Включает всех современных уховерток. В ископаемом состоянии известны начиная с юры (рис. 2.2). Помимо типичных уховерток к Neodermaptera относятся Diploglossata Saussure 1879 NomCirc. {syn.circ.: Dermodermaptera Verhoeff 1902c; nom.typ.: Hemimerus/fg [f:1895; g:1871]} (бескрылые эктопаразиты грызунов) и Plecodermaptera Crampton 1915 {nom.typ.: Arixenia/fg [fg:1909]} (бескрылые сожители рукокрылых), которых некоторые авторы не относили к отряду уховерток, а помещали в отдельные отряды.
* * *
KlugeIMGP2557.JPG)

Подробнее см. Н.Ю. Клюге 2020. «Систематика насекомых и принципы кладоэндезиса».