КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ
Глава I из книги «Систематика насекомых и принципы кладоэндезиса»

![]()
|
КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ Глава I из книги «Систематика насекомых и принципы кладоэндезиса» |
|||||||
|
|
|
fg | |||||
 I.1. Что такое
кладоэндезис
I.1. Что такое
кладоэндезис
![]() I.2. Понятия и
термины, связанные с биологической
эволюцией
I.2. Понятия и
термины, связанные с биологической
эволюцией
I.3. Биологический вид
![]() I.4.
Реконструирование
филогении
I.4.
Реконструирование
филогении
I.4.1. АНАЛИЗ ФИЛОГЕНИИ НА ОСНОВЕ СИНАПОМОРФИЙ
I.4.1.1. Термины апоморфия и плезиоморфия
I.4.1.2. Логика анализа филогении на основе синапоморфий
I.4.1.3. Взаимозависимость филогенетической теории и формулировки признаков
I.4.1.4. Область применения филогенетического анализа
I.4.1.5. Филогенетический анализ и эволюционный сценарий
I.4.1.6. Поиск апоморфий
I.4.2. ОШИБОЧНЫЕ ПРЕДСТАВЛЕНИЯ ОБ АНАЛИЗЕ ФИЛОГЕНИИ
I.4.2.1. Несостоятельность эмбриологического метода
I.4.2.2. Ограничения палеонтологического метода
I.4.2.3. Необязательность усложнения строения
I.4.2.4. Необязательность олигомеризации
I.4.2.5. Необязательность неспециализированности предка
I.4.2.6. Причины несостоятельности нумерических и матричных методов
I.4.2.6.1. Несостоятельность принципа экономности
I.4.2.6.2. Некорректность представления данных в виде матрицы
I.4.2.6.3. Фундаментальные недостатки нумерических методов в систематике
I.4.2.7. Подмена молекулярного метода анализа филогении
I.5.
Принципы классификации надвидовых
таксонов
I.6.
Принципы номенклатуры таксонов
I.4. Реконструирование филогении
Слово филогения, или филогенез (phylogenesis, от греческого φυλον – племя, и γενεσις – происхождение) было введено Э. Геккелем (Haeckel 1866) и означает то же, что Ч. Дарвин называл генеалогией.
Родственные связи разных таксонов (или филумов) можно графически изобразить в виде схемы, называемой филогенетическим древом, или кладограммой (cladogramma). Филогенетическое древо – это график из ветвящихся линий, где каждая линия означает существование во времени биологического вида с единым генофондом и разветвляется там, где происходит дивергенция и возникает репродуктивная изоляция (т.е. изоляция между генофондами); длина и направление линий бывают либо произвольными, либо привязаны к той или иной системе координат (можно по тем или иным осям откладывать геологическое время, накопление различий или другие параметры). Чаще рисуют более обобщенные филогенетические схемы, где каждая линия означает не отдельный вид, а филогенетическую ветвь, состоящую из множества видов; при этом подразумевают, что все разветвления этой ветви в данном масштабе как бы не различимы и поэтому могут быть изображены слитыми, то есть в виде сплошной полосы или линии. В любом случае каждое разветвление, изображенное на филогенетическом древе, означает дивергенцию – то есть момент, когда один биологический вид разделился на два вида, репродуктивно изолированных друг от друга. Следует предостеречь от чрезмерного увлечения аналогией между филогенетической схемой и деревом: филогенетическая схема похожа на дерево только тем, что ветвится; в отличие от настоящего дерева, филогенетическое древо не имеет ни корня, ни центрального ствола, ни толстых оснований ветвей.
Реконструкция филогении состоит в том, чтобы правильно выявить последовательность ветвлений филогенетического древа.
Выводы о тех или иных филогенетических связях и вообще о происхождении разных животных от общего предка делаются на основе того факта, что у разных животных имеются такие сходные признаки, которые нельзя объяснить ни случайным совпадением, ни какими либо универсальными законами биологии. Наличие такого рода сходных признаков свидетельствует, что эти признаки были унаследованы его обладателями от их общего предка. Тот факт, что разные животные, обладающие таким общим признаком, различаются по другим признакам, означает, что по крайней мере какие-то из этих животных также отличаются и от названного предка; таким образом, мы приходим к выводу о существовании эволюционных изменений.
Чтобы выяснить, какие именно эволюционные изменения и в какой именно последовательности происходили, необходимо проанализировать распределение общих признаков этих животных. Такой анализ называется филогенетическим, или кладистическим анализом (от греческого cladus – ветвь), поскольку он выявляет конкретные ветви филогенетического древа. Здесь под словосочетанием «филогенетический анализ» мы понимаем анализ филогении, основанный на научных гипотезах; однако тем же термином часто называют нумерический метод построения дендрограмм, в котором научные гипотезы преднамеренно игнорируются (см. ниже, I.4.2.6).
Основоположником филогенетического анализа считается немецкий энтомолог Вилли Хенниг (W. Hennig 1913–1976), опубликовавший в 1950 г. книгу «Grundzuge einer Theorie der phylogenetischen Systematik» (английский перевод этой книги «Phylogenetic systematics», ставший наиболее известным, вышел в 1966 г.). Однако теми же методами, которые были описаны Хеннигом, биологи пользовались до него уже не меньше полутора столетий. Например, еще во втором томе «Философии зоологии» Ж.-Б. Ламарка (Lamarck 1830) приведена схема филогении животных, которая могла быть построена только путем филогенетического анализа (хотя комментарии к этой схеме крайне скупы и не объясняют, каким методом пользовался автор). Особенно большое число работ по филогении появилось начиная со второй половины XIX века, когда благодаря работам Ч. Дарвина эволюционный подход в биологии приобрел всеобщее признание. При этом в работах большинства авторов эволюционные реконструкции фактически основаны на филогенетическом анализе, однако необходимые пояснения о методах реконструирования филогении в них, как правило, отсутствуют. Многие из этих авторов ясно представляли себе логику филогенетического анализа; другие авторы, интуитивно пользуясь этой логикой, на словах объясняли принципы реконструирования филогении как-то иначе. Из-за отсутствия четкой формулировки принципов анализа филогении у некоторых людей, читавших работы по филогении, складывалось превратное впечатление, что филогенетические схемы рисуются произвольно. Так что заслуга Хеннига состоит не в том, что он открыл новый метод, а в том, что он впервые последовательно описал методы реконструирования филогении и именно его публикации заставили последующих авторов ясно формулировать те принципы, которые они используют для своих филогенетических реконструкций.
Одной из заслуг Хеннига является также и то, что он ввел в употребление несколько важных терминов.
I.4.1.1. Термины апоморфия и плезиоморфия
Термин апоморфия (apomorphia, от греческого απο- – от), или апоморфный признак, означает прогрессивный (в исходном значении этого слова, от латинского pro- – вперед и gressus – движение), т.е. продвинутый (или производный, вторичный) признак. Термин плезиоморфия (plesiomorphia, от греческого πλησιος – близкий), или плезиоморфный признак, означает примитивный (в исходном значении этого слова, от латинского primus, primitivus – первый, первичный) признак (называемый также исходным, анцестральным, предковым или генерализованным признаком). Плезиоморфия и апоморфия – это два альтернативных состояния признака, соответственно, до и после произошедшего с признаком эволюционного изменения. Эти понятия являются относительными: если эволюция идет от состояния A к состоянию B и далее к C, то B является апоморфией по отношению к A, но плезиоморфией по отношению к C. Представим себе, что в ходе эволюции форма какой-то части тела животного изменилась от круглой к квадратной и далее к треугольной; в этом случае квадратная форма будет апоморфией по отношению к круглой и плезиоморфией по отношению к треугольной:
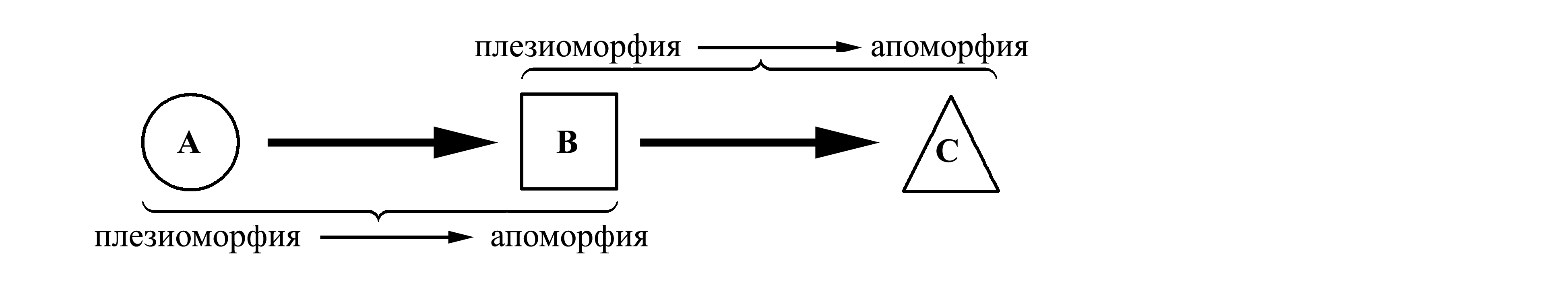
Термины «прогрессивный» и «примитивный» нередко связывают с понятиями «более совершенный» и «менее совершенный» или «более сложный» и «более простой», что может внести путаницу. Термины «апоморфный» и «плезиоморфный» имеют только одно значение – направление эволюционного изменения данного признака в данном случае, что делает эти термины более строгими.
Апоморфия, отличающая данный таксон от всех прочих, называется его аутапоморфией, или автапоморфией (autapomorphia), а апоморфия, общая для нескольких таксонов, называется их синапоморфией (synapomorphia). Плезиоморфия, общая для нескольких таксонов, называется их симплезиоморфией (symplesiomorphia).
I.4.1.2. Логика анализа филогении на основе синапоморфий
Филогенетический анализ базируется на предположениях о неповторимости и о необратимости эволюционных преобразований [последнее известно как закон Долло (Dollo 1893)]. Эти предположения, в свою очередь, основываются на соображениях вероятности: вероятность того, что одно и то же изменение в генотипе может произойти независимо несколько раз, настолько мала, что ее можно считать равной нулю. Таким образом, предполагается, что каждая апоморфия возникла один раз и унаследована всеми потомками того вида, у которого она возникла. Следовательно, если у разных видов имеется один и тот же апоморфный признак (т.е. синапоморфия), это свидетельствует о том, что все эти виды унаследовали этот признак от своего общего предка и, следовательно, эти виды образуют единую филогенетическую ветвь; в то же время наличие у разных видов общих плезиоморфных признаков (т.е. симплезиоморфий) ни о чем не свидетельствует. Таким образом, при анализе филогении мы анализируем синапоморфнное сходство, игнорируя симплезиоморфное сходство.
Следует обратить внимание на то, что для доказательства филогенетических связей животных, так же как и для доказательства существования биологической эволюции вообще, мы используем именно сходства между животными, а не различия. Хотя различия между разными видами животных возникли в ходе эволюции, они не достаточны для доказательства эволюции, поскольку различия между двумя объектами могут быть обеспечены не только изменениями при превращении одного объекта в другой, но и независимым происхождением этих объектов.
Для того чтобы доказать, что некие виды животных образуют единую филогенетическую ветвь, необходимо выявить у них синапоморфию, которая при этом являлись бы аутапоморфией для всей этой ветви. Для этого необходимо установить, какой из альтернативных признаков является апоморфным.
Для выявления полярности признака (т.е. для выяснения, какое состояние признака является плезиоморфным, а какое апоморфным) используется принцип внешней группы, который выражается в следующем. Если мы анализируем филогенетические связи внутри определенной филогенетической ветви и наблюдаем у ее представителей два альтернативных состояния признака, одно из которых является плезиоморфией, а другое апоморфнией, то за пределами данной филогенетической ветви (т.е. во внешней группе) может быть встречена та же самая плезиоморфия, но не может быть встречена та же самая апоморфия. Это утверждение вытекает из предположения, что данная апоморфия возникла в результате однократного неповторимого эволюционного процесса, произошедшего внутри анализируемой филогенетической ветви, и поэтому за пределами этой ветви оказаться не может. В отличие от апоморфии, плезиоморфия была унаследована от какого-то более раннего предкового вида, который может быть предком не только для анализируемой нами филогенетической ветви, но и для других филогенетических ветвей, которые по отношению к рассматриваемой нами ветви являются частью внешней группы. Некоторые авторы полагают, что для филогенетического анализа достаточно в качестве внешней группы использовать признаки одного вида; это совершенно неверное толкование «принципа внешней группы», поскольку по одному или нескольким произвольно выбранным видам никак нельзя судить о том, встречается ли данный признак во всей внешней группе. В действительности внешней группой является не один вид, и не какой-либо заведомо ограниченный набор видов, а вся совокупность живых организмов за пределами анализируемой филогенетической ветви. Поскольку на практике исследователь не может учесть все живые организмы (хотя бы потому, что не все они открыты), приходится ограничиваться известными данными.
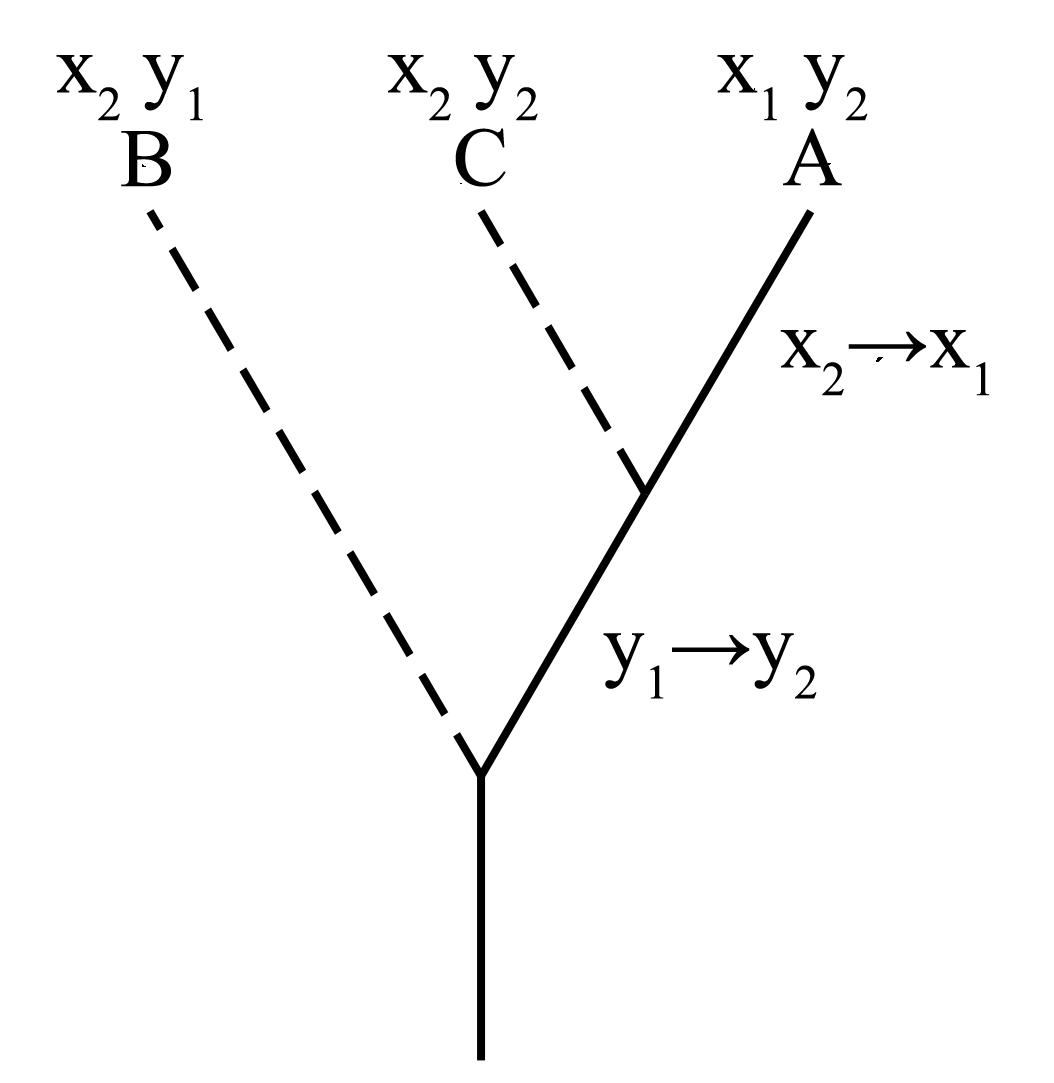
Задача о трех таксонах. Возьмем наиболее простую формальную задачу – выявление филогенетических отношений между тремя таксонами A, B и C. Если считать, что эволюция состоит из последовательных дивергенций, а разделение одного предка сразу на три или более ветвей маловероятно, то между тремя таксонами теоретически возможны три варианта филогенетических взаимоотношений:
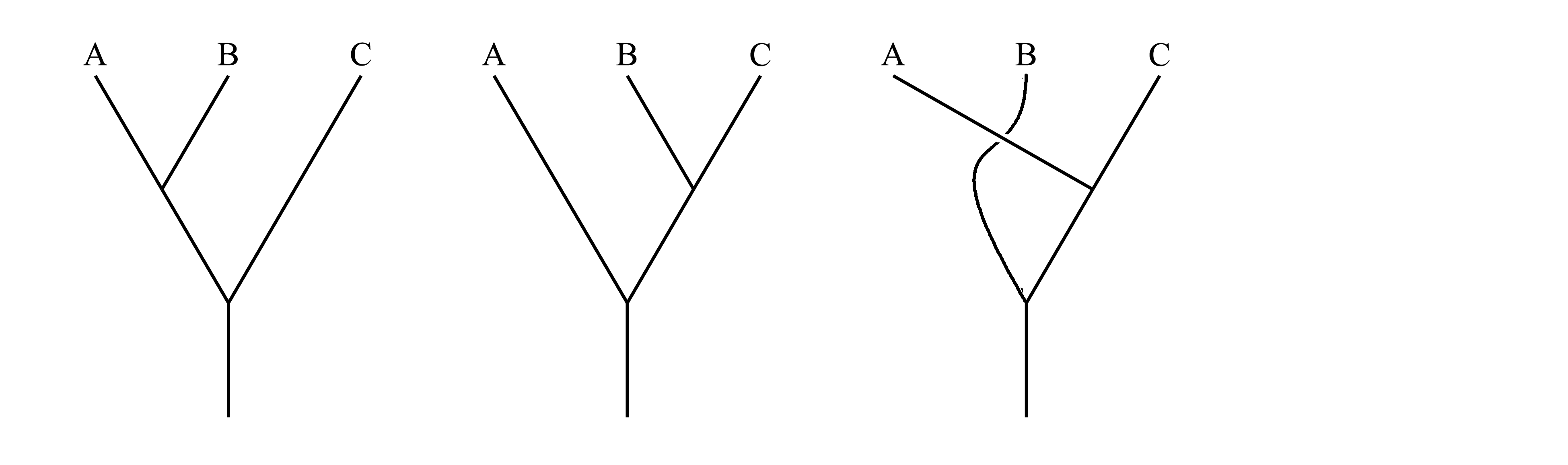
Для того, чтобы выяснить, какой из трех вариантов верный, необходимо проанализировать признаки этих трех таксонов. Обозначим маленькими буквами a, a', b и b' признаки, где a и a' являются двумя альтернативными состояниями одной детали строения, а признаки b и b' являются двумя альтернативными состояниями другой детали строения. Предположим, что признаки a и b являются плезиоморфиями, а признаки a' и b' – апоморфиями, то есть их эволюционные преобразования можно записать так: a→a' и b→b'. В нашем примере таксон A имеет признаки a и b, таксон B имеет признаки a и b', таксон C имеет признаки a' и b':

Таким образом, таксоны A и B имеют общий признак a, тогда как таксоны B и C имеют общий признак b'. Поскольку признак a является симплезиоморфией, а признак b' является синапоморфией, мы игнорируем наличие симплезиоморфии a у таксонов A и B, а на основании наличия синапоморфии b' у таксонов B и C делаем вывод, что таксоны B и C образуют единую филогенетическую ветвь, к которой не относится таксон A (сплошными линиями показаны ветви, существование которых доказано в этом примере):

Мы получили обоснованную реконструкцию филогении, в которой каждое из эволюционных преобразований (a→a' и b→b') произошло однократно.
Здесь важно отметить, что для реконструирования древа, ветви которого включают более чем по одному виду, необходимо не только выявить синапоморфии между ветвями, но и найти аутапоморфии каждой ветви. Если для какого-то таксона, состоящего из некоторого числа репродуктивно изолированных биологических видов не выявлено ни одной аутапоморфии, мы не имеем оснований изображать этот таксон в виде филогенетической ветви. Так что на вышеприведенной схеме помимо апоморфий a' (для ветви C) и b' (для ветви B+C) должны быть указаны также аутапоморфии для ветви A и для ветви B; если такие аутапоморфии неизвестны, то неизвестно, являются ли таксоны A и B особыми филогенетическими ветвями, или таксон A идентичен общему предку A, B и C, а таксон B идентичен общему предку B и C. Здесь пунктирными линиями условно изображены те ветви, существование которых не доказывается в данном примере.
Для того, чтобы в рассматриваемом примере определить полярность признаков a и a', b и b', необходимо обратиться к внешней группе, т.е. к филогенетическим ветвям за пределами ветви A+B+C. На схеме, приведенной ниже, у таксонов D, E, F и G признаки, отличные от признаков a, a', b и b', показаны многоточиями (...). У некоторых из этих таксонов имеется признак a, у некоторых имеется признак b, но ни у кого не встречается ни a', ни b'. Из этого следует, что признаки a и b являются плезиоморфиями, унаследованными от предка, общего для всех этих ветвей, а признаки a' и b' – апоморфии, возникшие внутри ветви A+B+C.
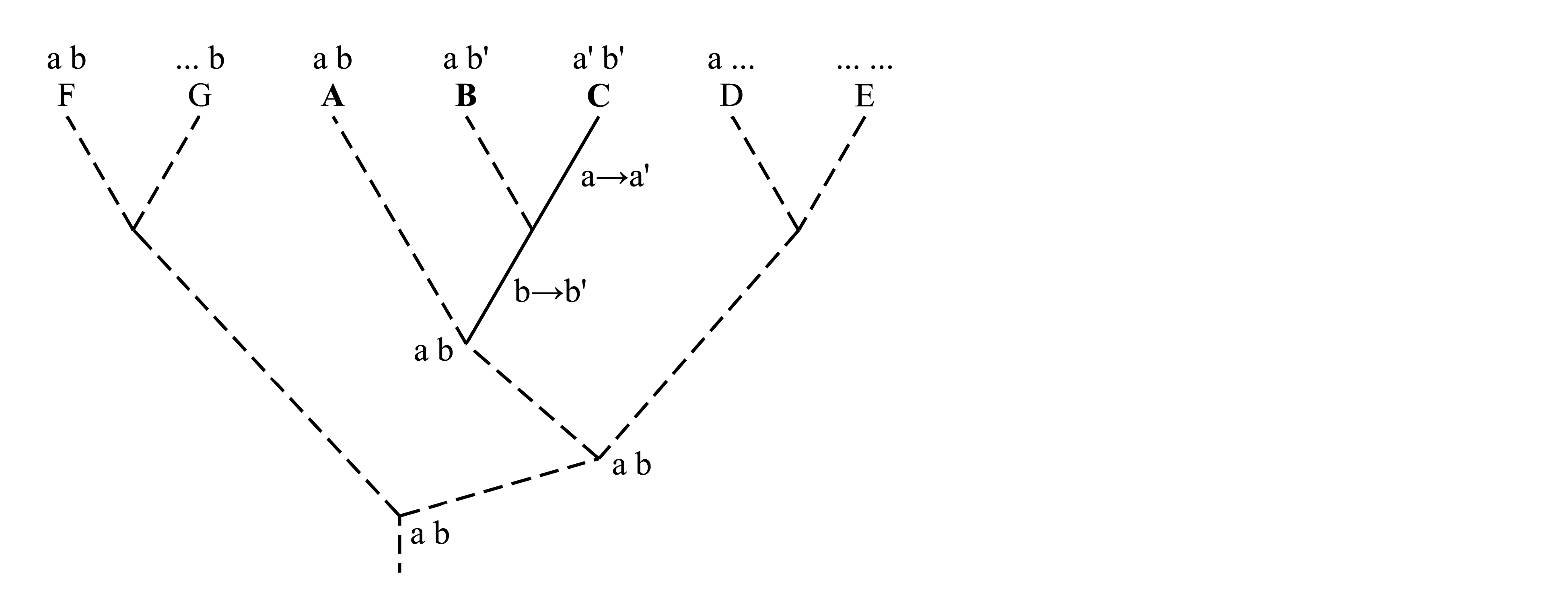
Однако приведенных здесь фактов недостаточно для доказательства именно такой полярности признаков a, a', b и b' и недостаточно для доказательства именно таких филогенетических взаимоотношений между таксонами A, B и C. Основываясь на том же распределении признаков, мы можем допустить совершенно иную их полярность и иное ветвление филогенетического древа:
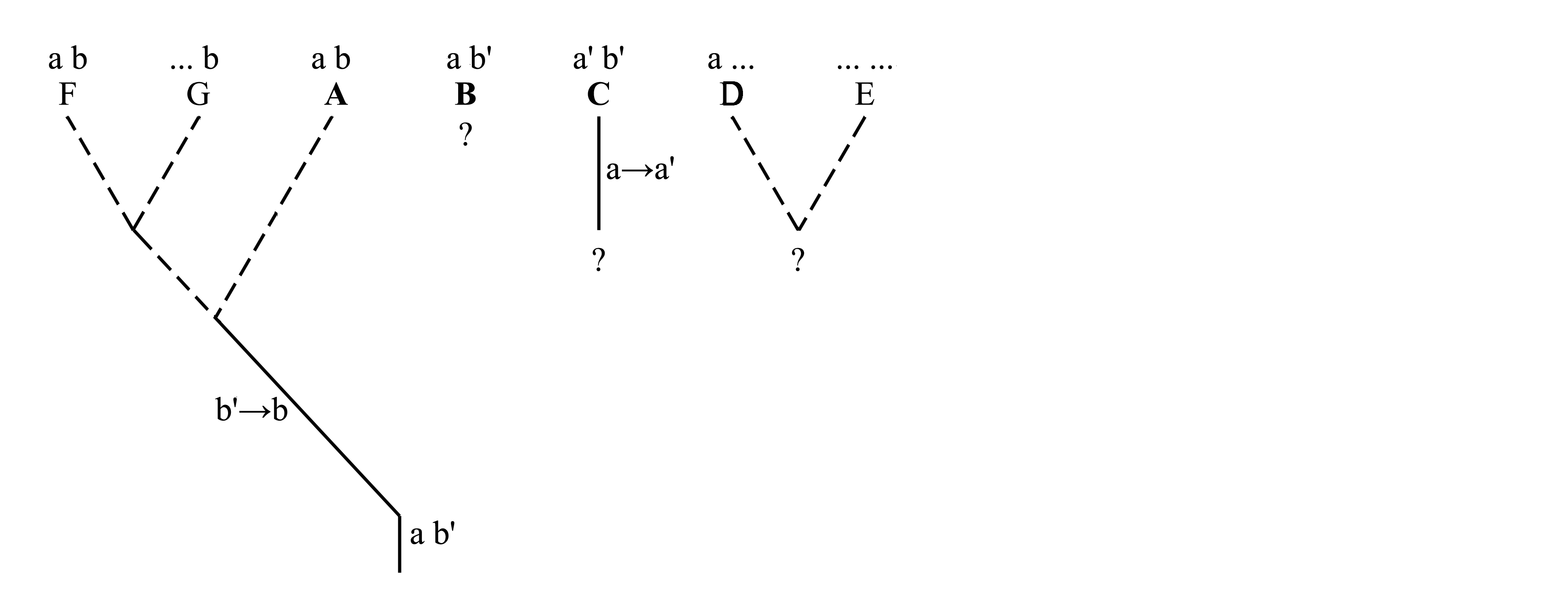
Здесь мы предположили, что апоморфией является не b', а b, и в этом случае получилось, что таксон A имеет синапоморфию с ветвью F+G, а родство между таксонами B и C оказалось недоказанным. Такое предположение оказалось возможным, потому что мы не доказали, что таксоны D, E, F и G относятся к внешней группе. Чтобы это доказать, необходимо доказать существование филогенетической ветви A+B+C. Единственный способ доказать существование филогенетической ветви – это найти ее аутапоморфию, т.е. синапоморфию ее представителей. Анализируя филогенетические взаимоотношения между таксонами A, B и C, мы использовали только признаки a–a' и b–b' и проигнорировали те признаки, которые идентичны у всех трех таксонов A, B и C. Теперь добавим признак, имеющий плезиоморфное состояние c и апоморфное состояние c':

Теперь существование ветви A+B+C доказано наличием аутапоморфии c'. Тем самым доказано, что D, E, F и G относятся к внешней группе, поэтому наличие в этих таксонах признака b свидетельствует о том, что это плезиоморфия; следовательно, b' является апоморфией и поэтому наличие этого признака у таксонов B и C служит доказательством их родства. В этой цепи рассуждений необходимо еще доказать, что признак c' является апоморфией, т.е. что его полярность выглядит как c→c'. Для этого нужно обратиться к внешней группе, т.е. к филогенетическим ветвям за пределами ветви A+B+C+D+E+F+G и найти апоморфию, доказывающую существование этой ветви.
Таким образом, процесс филогенетического анализа не имеет начала и конца. Невозможно, имея объективные факты о признаках животных и применяя правила филогенетического анализа, последовательно шаг за шагом реконструировать филогенетическое древо, не делая при этом ошибок. Реконструирование филогении с помощью филогенетического анализа представляет собой непрерывный процесс поиска непротиворечивой теории и исправления ранее сделанных ошибок. Как показано ниже, признаки, на которых основывается реконструирование филогении, также не являются исходными данными, а сами в определенной мере зависят от результатов реконструирования филогении.
I.4.1.3. Взаимозависимость филогенетической теории и формулировки признаков
На первый взгляд кажется, что признаки животных являются объективно существующими и исследователям остается их лишь констатировать. Однако на самом деле признаков как таковых в природе не существует, а существует лишь строение организмов. Признаки формулируются нами на основании объективно имеющегося в природе строения организмов, преломленного нашим восприятием. Наше же восприятие признаков зависит от нашего представления о филогении этих животных.
Например, на такой простой вопрос для младших школьников, сколько ног у жука и у паука, не может быть единого правильного ответа (табл. I.2).
![]() Таблица
I.2. Число ног у некоторых членистоногих, посчитанное разными способами.
Таблица
I.2. Число ног у некоторых членистоногих, посчитанное разными способами.
|
Способы |
Некоторые группы членистоногих |
|||||
|
Hexapoda |
личинки
|
личинки
|
Arachnida |
Xiphosura |
Decapoda |
|
|
Способ
1: |
3 |
3 |
3 |
4 |
5 |
5 |
|
Способ
2: |
||||||
|
I |
|
|||||
|
II |
+ |
|||||
|
III |
+ |
+ |
+ |
|||
|
IV |
+ |
+ |
+ |
|||
|
V |
+ |
+ |
+ |
+ |
|
|
|
VI |
+ |
+ |
+ |
+ |
||
|
VII |
+ |
+ |
||||
|
VIII |
+ |
|||||
|
IX |
+ |
|||||
|
X |
+ |
|||||
|
XI |
+ |
|||||
|
XII |
+ |
|||||
Если просто пересчитать ходильные ноги, то получатся числа, показанные в строке «Способ 1»; это лишь один из множества возможных способов подсчета: здесь, например, в качестве ног посчитаны клешни раков (Decapoda), но не посчитаны такие же на вид клешни скорпионов (т.е. педипальпы паукообразных – Arachnida). При таком подсчете можно усмотреть сходство между Hexapoda, личинками Diplopoda и личинками Acari (по 3 пары ног) и между Xiphosura и Decapoda (по 5 пар ног); во многих классификациях XIX века Xiphosura и Decapoda помещались в таксон Crustacea auct.
В настоящее время этот же признак формулируют иначе («Способ 2»). При такой формулировке оказывается, что Xiphosura и Decapoda не имеют в этом отношении ничего общего, так же как личинки Acari и Hexapoda; зато обнаруживается синапоморфия между Arachnida и Xiphosura, на основании чего они объединяются в таксон Chelicerata (см. главу IV). Имеется также общий признак Hexapoda и Diplopoda, который можно истолковать как синапоморфию. Показанная здесь интерпретация расположения ног основывается на целой серии принятых нами допущений. А) Мы считаем, что у членистоногих ноги каждой определенной пары связаны с определенным сегментом; однако с этим утверждением не согласны некоторые современные палеонтологи; это утверждение верно именно для членистоногих, но неверно, например, для позвоночных. Б) Мы считаем, что гомологичным для всех членистоногих является сегмент, находящийся непосредственно позади ротового отверстия (т.е. первый посторальный сегмент), и что наиболее достоверно этот сегмент может быть выявлен по его иннервации [см. главу IV: Metameria (2)]; не все зоологи согласны с этим утверждением: по мнению некоторых авторов иннервация не является надежным указанием на первый посторальный сегмент, а по мнению других, ротовое отверстие способно в ходе эволюции менять свое положение, так что первый посторальный сегмент не гомологичен у разных членистоногих (см. там же). В) Мы отсчитываем сегменты, начиная с переднего, и считаем, что в этом случае сегменты с равными номерами должны быть гомологичны; в то же время очевидно, что, если отсчитывать сегменты с заднего конца тела, этого не будет. Однако в некоторых случаях наблюдается обратная картина: например, у всех Chilopoda два последних сегмента (прегенитальный и генитальный) явно гомологичны; в то же время общее число туловищных сегментов у разных хилопод различное, так что, если отсчитывать генитальный и прегенитальный сегменты спереди, их номера будут различаться у разных видов и даже особей. Г) Мы считаем, что придатки первого посторального сегмента у Hexapoda и Diplopoda исчезли вторично [см. главу IV: Atelocerata (1)]; однако по мнению некоторых авторов, этого сегмента никогда не было, и в таком случае вся схема должна выглядеть иначе. Д) Мы считаем, что хилярий Diplopoda образован придатками только III посторального сегмента, а коллюм является IV сегментом и лишен конечностей; это также является предметом дискуссий [см. главу V: Collifera (1)–(2)]. Е) Мы считаем, что три пары ног гомологичны у всех Diplopoda; это отнюдь не очевидно и потребовало специальных доказательств [см. главу V: Diplopoda (3)]. Ж) Мы считаем, что у Diplopoda сегменты, обозначенные здесь как V–VII, являются гаплосегментами; это, возможно, неверно (см. там же). З) Мы считаем, что эти же сегменты у Hexapoda являются гаплосегментами, по поводу чего также существуют сомнения [см. главу VI: Hexapoda (1)]. И) Мы считаем, что суперлингвы Hexapoda не являются конечностями, как думали некоторые авторы [см. главу IV: Mandibulata (3)].
Этот список допущений и разногласий можно продолжить. В зависимости от того, какие из допущений мы принимаем, а какие нет, мы усматриваем или не усматриваем у тех или иных таксонов общие признаки, в том числе синапоморфии. Таким образом, от этих допущений зависит результат филогенетического анализа. В то же время каждое из этих допущений сделано на основании определенного представления о филогении: например, первое допущение (А) основано на представлении о филогенетическом единстве Metameria и каждой группы членистоногих, обладающей определенным набором конечностей. Эти представления о филогении, в свою очередь, основаны на филогенетическом анализе каких-то иных признаков, интерпретация которых так же не является однозначной и зависит от представлений о филогении, и так далее, до бесконечности.
На этом простом примере мы видим, что представление о филогении зависит от формулировки признаков, а формулировка признаков зависит от представлений о филогении. В связи с этим существует опасность попасть в порочный круг доказательств, когда неверное представление о филогении и неверная формулировка признаков доказывают друг друга. Порочный круг возникает тогда, когда набор аргументов замкнут, поэтому, чтобы разорвать порочный круг, надо вводить новые аргументы. При реконструировании филогении можно ввести новые аргументы двумя способами: 1) расширить круг изучаемых видов; это можно сделать, открывая новые, неизвестные науке виды (для многих групп насекомых это вполне реальный путь исследований, поскольку ежегодно открываются тысячи новых видов); 2) расширить круг анализируемых признаков; этого можно достичь, привлекая новые методики и изучая те структуры, которые прежде были мало исследованы или не исследованы.
I.4.1.4. Область применения филогенетического анализа
В филогенетическом анализе можно и нужно использовать любые наследуемые признаки организмов – анатомические, гистологические, цитологические, биохимические, физиологические, этологические и прочие, относящиеся к любым стадиям онтогенеза.
Филогенетический анализ в его классической форме предназначен для реконструирования филогении, основанной на дивергенциях, когда филогенетическая схема имеет вид иерархически ветвящегося древа, но не сети. Однако в некоторых случаях к обычным эволюционным механизмам дивергенции примешиваются такие факторы, как гибридизация или параллельный перенос генов, в результате чего гены передаются не только от единого предка к его дивергирующим потомкам, но и другим способом, так что филогенетическое древо приобретает сетчатый вид.
В принципе филогенетический анализ может быть применен к таксонам, имеющим ранг не ниже видового. Между внутривидовыми таксонами нет репродуктивной изоляции, так что их родственные отношения имеют вид сети, а не филогенетического древа, и не могут быть выявлены филогенетическим анализом.
Многие исследователи используют филогенетический анализ для реконструкции филогенетических отношений между отдельными видами. Теоретически это правомерно, однако практически, вероятно, малоприемлемо, так как анализ на уровне видов не позволяет выявить и исключить из анализа те признаки, которые способны существовать в генотипе в скрытом виде и появляться в фенотипе как независимо возникающие гомологичные признаки.
Помимо синапоморфий (т.е. признаков, унаследованных от общего предка), существуют гомологичные, но независимо возникающие признаки. Связано это с тем, что от общего предка может быть унаследован не фенотипический признак, а только кодирующие его гены; при последующих эволюционных изменениях такой скрытый признак может появляться в фенотипе, и это может происходить независимо в разных филогенетических ветвях. Появляющийся независимо гомологичный признак может быть ошибочно принят за синапоморфию, и тогда филогенетический анализ приводит к ложным результатам. Чем ближе родство таксонов, тем более вероятно, что они содержат скрытые гомологичные признаки. Если каждый из анализируемых таксонов содержит много разнообразных видов, можно ожидать, что общие для каждого таксона скрытые признаки проявляются в фенотипе у каких-то из его видов. Тогда независимо появляющиеся признаки оказываются выявленными, и их можно исключить из филогенетического анализа. Поэтому чем выше ранг и чем больше разнообразие каждого из таксонов, включаемых в филогенетический анализ, тем больше достоверность этого анализа.
Наименее достоверным является филогенетический анализ для таксонов видового ранга (наименьшего ранга, для которого вообще теоретически возможен филогенетический анализ).
I.4.1.5. Филогенетический анализ и эволюционный сценарий
До того, как был сформулирован принцип внешней группы, высказывались различные иные соображения о том, как следует определять направление эволюции. В действительности единственным обоснованным методом определения полярности признака является принцип внешней группы, который состоит в том, что плезиоморфный признак, будучи унаследованным от более раннего предка, может встречаться за пределами рассматриваемой нами филогенетической ветви, а апоморфный признак, возникший в пределах данной ветви, за ее пределами отсутствует (см. I.4.1.2).
Например, в пределах таксона Hexapoda у одних представителей антенны имеют мышцы только в первом членике, у других – во всех члениках, кроме последнего. Если основываться только на строении антенн как таковом, можно было бы сколь угодно долго и бесплодно рассуждать о том, что здесь первично, а что вторично – более простые антенны с ограниченной подвижностью или более сложные антенны, способные совершать разнообразные движения. Рассуждения о том, что эволюция якобы обязательно должна быть куда-то направлена – в сторону усложнения или, наоборот, в сторону упрощения строения; в сторону увеличения адаптации к окружающей среде или, наоборот, в сторону уменьшения адаптации; в сторону большей перспективности для дальнейшей эволюции, или наоборот, в сторону эволюционного тупика – подобные рассуждения в данном случае не дают правильного ответа, поскольку на самом деле эволюционные изменения бывают бесконечно разнообразными и могут иметь любое направление. Принцип внешней группы дает однозначный ответ на вопрос о первичном строении антенн у Hexapoda: плезиоморфией (т.е. первичным состоянием) является наличие мышц во всех члениках антенны, кроме последнего, а апоморфией – наличие мышц только в первом членике: за пределами Hexapoda многочлениковые антенны с мышцами только в первом членике не встречаются, тогда как антенны с мышцами во многих члениках имеются у самых различных членистоногих. Отсюда следует вывод, что строение антенн с мышцами только в первом членике является апоморфией, возникшей в пределах таксона Hexapoda, так что этот признак является аутапоморфией подчиненного таксона Amyocerata в составе Hexapoda (см. главу VI).
Некоторые авторы полагают, что для реконструирования филогении достаточно построить эволюционный сценарий, т.е. описать предполагаемый путь эволюции с указанием возможных причин каждого эволюционного изменения. При этом эволюционные изменения животных рассматриваются в сочетании с внешними условиями, на фоне которых происходили эти изменения. Для объяснения направления действия естественного отбора привлекаются данные о сменах климатов и ландшафтов в соответствующие геологические эпохи. В действительности же каждый такой сценарий является лишь одной из многих возможных гипотез.
Например, предлагались сценарии, описывающие эволюционный путь от кольчатых червей – к онихофорам – к многоножкам – к насекомым. При этом предполагается, что многощетинковые кольчатые черви (Polychaeta), исходно обитающие в морях, вышли на сушу; в результате приспособления к жизни на поверхности суши их торчащие в стороны многочисленные параподии сместились на вентральную сторону тела и превратились в нечленистые ножки, которые характерны для Onychophora; дальнейшая эволюция этих животных происходила как совершенствование приспособления к жизни на суше, их ноги приобрели членистость, передние пары конечностей вошли в состав ротового аппарата, появились другие особенности, характерные для членистоногих, в результате чего Onychophora дали начало многоножкам (Myriapoda) и далее – насекомым (Hexapoda).
В этом сценарии не нашлось места для других членистоногих – хелицеровых (Chelicerophora), ракообразных (Eucrustacea) и трилобитообразных (Trilobitomorpha), поэтому приверженцы вышеописанного сценария считают, что эти группы животных произошли от кольчатых червей независимо от Myriapoda и Hexapoda. В соответствии с этим они предлагают эволюционные сценарии независимого происхождения хелицеровых и ракообразных от кольчатых червей. При этом считают, что конечности ракообразных, произошедшие непосредственно из параподий Polychaeta, исходно были нерасчлененными (плавательные листовидные ножки – филлоподии, характерные для жаброногов – Branchiopoda), и расчленились в ходе эволюции ракообразных. На основании таких эволюционных сценариев был сделан вывод о полифилии членистоногих (см. главу IV: Gnathopoda: «Классификации» II).
Однако существует и совершенно иная теория происхождения членистоногих, согласно которой членистоногие представляют собой голофилетический таксон (см. главу IV: Euarthropoda). Этой теории соответствует не менее убедительный эволюционный сценарий, чем предыдущий: Polychaeta дали начало трилобитообразным, при этом их конечности расчленились не в связи с наземным образом жизни, а в связи с обитанием на дне моря; членистые конечности, унаследованные от трилобитообразных, сохранились и у ракообразных (причем у Branchiopoda конечности некоторых пар вторично утратили расчленение в связи с превращением в плавательные органы); передние пары конечностей ракообразных видоизменились, войдя в состав ротового аппарата; далее следует выход на сушу и появление особенностей, характерных для Myriapoda и Hexapoda.
Две приведенные гипотезы являются взаимоисключающими. Ясно, что одни и те же насекомые не могли возникнуть двумя разными путями, и верна лишь одна из этих гипотез. Обе гипотезы кажутся правдоподобными, пока мы ограничиваемся построениями лишь эволюционных сценариев. Помимо названных двух, предлагалось много других взаимоисключающих эволюционных сценариев о происхождении насекомых, в каждом из которых каждый отдельный этап выглядит правдоподобно.
Как видно, одного только построения эволюционного сценария недостаточно для доказательства филогенетической гипотезы.
Аргументированное представление о филогении можно получить лишь с помощью филогенетического анализа, т.е. реконструкции филогении на основе апоморфий. Для этого надо прежде всего изобразить рассматриваемые гипотезы не в виде эволюционных рядов, а в виде кладограмм: эволюционный ряд состоит в основном из гипотетических форм, не поддающихся прямому изучению, а в кладограмме каждая ветвь завершается современным таксоном, все свойства которого можно изучить.
Вместо эволюционного ряда «Polychaeta → Onychophora → Myriapoda → Hexapoda» следует нарисовать следующую кладограмму:

Вместо эволюционного ряда «Polychaeta → Trilobitomorpha → Eucrustacea → Myriapoda → Hexapoda» следует нарисовать следующую кладограмму:
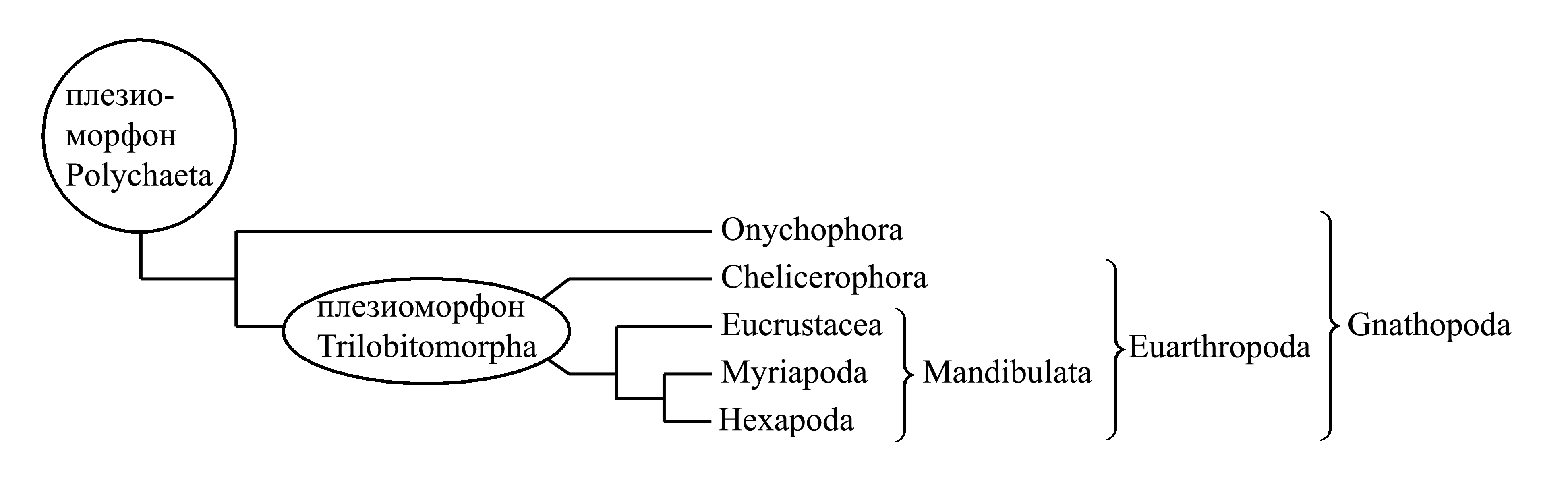
Доказательством первой из этих гипотез, и одновременно опровержением второй, могли бы быть синапоморфии таксонов Onychophora, Myriapoda и Hexapoda, которые в таком случае являлись бы аутапоморфиями таксона Ceratophora, поскольку в первой теории таксон Ceratophora представляется как особая филогенетическая ветвь. Во второй же гипотезе Onychophora отделяются от общего предка Myriapoda и Hexapoda раньше, чем тот отделяется от Eucrustacea, Trilobitomorpha и Chelicerophora. Следовательно, согласно второй гипотезе, у Onychophora, Myriapoda и Hexapoda не может быть таких синапоморфий, которые отсутствовали бы у Eucrustacea, Trilobitomorpha и Chelicerophora. В действительности синапоморфий Onychophora, Myriapoda и Hexapoda (то есть аутапоморфий Ceratophora) до сих пор не обнаружено.
Доказательством второй из этих гипотез, и одновременно опровержением первой, являются, во-первых, аутапоморфии Gnathopoda; во-вторых, синапоморфии Chelicerophora, Trilobitomorpha, Eucrustacea, Myriapoda и Hexapoda – т.е. аутапоморфии Euarthropoda; в-третьих, синапоморфии Eucrustacea, Myriapoda и Hexapoda – т.е. аутапоморфии Mandibulata. Эти комплексы апоморфий существуют на самом деле (см. главу IV). Гипотеза о голофилии членистоногих (вторая кладограмма) объясняет все эти случаи присутствия единых признаков у разных животных как результат наследования их от общего предка – т.е. как синапоморфии. В гипотезе же о полифилии членистоногих (первая кладограмма) существование филогенетических ветвей, соответствующих таксонам Gnathopoda, Euarthropoda, и Mandibulata, не признается и, следовательно, предполагается, что их общие признаки не являются синапоморфиями. Поэтому приверженцы теории о полифилии членистоногих пытались объяснить наличие этих признаков у разных животных конвергенцией, возникшей как результат приспособления к одинаковым условиям. Эти объяснения выглядят весьма натянуто, так как трудно придумать, какие одинаковые условия имеются в морских глубинах и на поверхности суши, где обитают членистоногие, так же как трудно придумать, какое адаптивное значение могут иметь такие признаки, как утрата целома или наличие дополнительной светопреломляющей линзы в каждом омматидии фасеточного глаза. Таким образом, при рассмотрении двух взаимоисключающих теорий филогенетический анализ позволяет сделать выбор в пользу теории о голофилии членистоногих: эта теория обосновывается конкретными апоморфиями (хотя и оспариваемыми оппонентами), тогда как альтернативная ей теория о полифилии членистоногих никак не обосновывается.
I.4.1.6. Поиск апоморфий
Главной трудностью филогенетического анализа является поиск апоморфий, доказывающих существование тех или иных филогенетических ветвей (или, что то же самое – голофилию тех или иных таксонов – см. I.5.1). Такие апоморфии могут содержаться в строении любых органов, а могут отсутствовать вовсе. Если исследователь ставит перед собой задачу найти синапоморфию таксонов A и B (или, что то же самое, аутапоморфию таксона, объединяющего A и B), он может искать ее где угодно – начиная от внешней формы или цвета животного, до ультратонкого строения любой части организма или до последовательности нуклеотидов в любом месте генома. При этом успех поиска не гарантирован. Если апоморфия не найдена, это может означать либо то, что гипотеза о сестринском родстве таксонов A и B неверна, либо то, что она верна, но в строении изучаемых структур нет ни одной синапоморфии A и B, либо то, что искомые апоморфии существуют, но остались не найденными. В ходе поиска апоморфий бывает неясно, до каких пор следует продолжать эти попытки, и можно потратить много усилий без nрезультата.
Рациональнее вести поиск апоморфий с другой стороны – не от гипотезы о филогении к поиску апоморфий, а от апоморфий к поиску их места в иерархической филогенетической системе животных. Этот метод и используется в кладоэндезисе. Он состоит в том, чтобы (1) записать каждый известный признак как аутапоморфию определенного голофилетического таксона и (2) от каждой такой аутапоморфии дать ссылку на исходную для нее аутапоморфию вышестоящего таксона. Логика этого метода состоит в том, что каждый признак, о котором есть основания предполагать, что он мог возникнуть в эволюции лишь единожды, является аутапоморфией какой-то одной и только одной филогенетической ветви. Если мы уже знаем о существовании такого признака, остается только найти ту филогенетическую ветвь, аутапоморфией которой данный признак является. В отличие от подбора аутапоморфий для ветви, подбор ветви для аутапоморфии в принципе всегда должен заканчиваться успехом: в то время как не у всякой филогенетической ветви есть аутапоморфии, всякая апоморфия является аутапоморфией какой-то филогенетической ветви. Чтобы зафиксировать результат поиска, необходимо придать филогенетической ветви статус таксона (такой таксон является голофилетическим – см. I.5.1) и дать этому таксону название. Имея название голофилетического таксона, можно записать под ним все аутапоморфии, выявленные для этого таксона. Если такой таксон был известен ранее, следует лишь дополнить его характеристику новыми аутапоморфиями; если такого таксона до сих пор не существовало, его следует установить и снабдить названием.
Например, если в качестве аутапоморфии маленького специализированного таксон Strepsiptera (веерокрылые) указана утрата анального отверстия [см. Strepsiptera (4)], то следует дать ссылку на тот вышестоящий таксон, аутапоморфией которого является приобретение анального отверстия. Этим таксоном оказывается малоизвестный таксон Haemataria, который объединяет большинство многоклеточных животных и характеризуется наличием сквозного кишечника [см. Haemataria (1)]. В некоторых современных классификациях нет места для Haemataria; авторы этих классификаций не обсуждают причину отказа от этого таксона, попросту игнорируя его. Необходимость делать ссылки на таксон, обладающий сквозным кишечником, заставляет либо признать реальность таксона Haemataria и отказаться от противоречащих этому гипотез о филогении многоклеточных животных, либо высказать гипотезу о многократном появлении анального отверстия, обосновать ее и установить таксоны, характеризующиеся независимым появлением анального отверстия.
До недавнего времени главная трудность состояла в том, чтобы снабдить названиями все таксоны, необходимые для филогенетического анализа. Поскольку традиционная типифицированная ранговая номенклатура позволяет снабдить названиями лишь ограниченное число таксонов (см. I.6.3), а нетипифицированные названия до недавнего времени использовались хаотично, многие филогенетические ветви оставались без названий. Чтобы выйти из положения, их обозначали порядковыми номерами или иными произвольными знаками. Каждая такая нумерация ветвей может быть использована только в пределах одной книги или статьи, где из-за ограниченного объема текста автор бывает вынужден указывать не все аутапоморфии. В других публикациях использовались иные обозначения для тех же филогенетических ветвей, и указывались иные апоморфии, в результате чего суммирование и накопление знаний об апоморфиях этих ветвей было затруднено. Создание рациональных универсальных безранговых номенклатур (см. I.6.4 и I.6.5) позволило решить эту проблему и снабдить однозначными названиями все выявляемые филогенетические ветви и вообще все таксоны (включая плезиоморфоны), которые приходится создавать в процессе реконструирования филогении.
Чтобы записать аутапоморфию таксона, обычно приходится упоминать либо более общую особенность строения, либо предковое состояние данной структуры. Каждое такое упоминание следует снабдить ссылкой на соответствующую аутапоморфию конкретного вышестоящего голофилетического таксона (что теперь стало возможным, благодаря наличию у каждого таксона собственного названия). Необходимость делать такие ссылки заставляет автора критически пересматривать распределение известных признаков.
I.4.2. ОШИБОЧНЫЕ ПРЕДСТАВЛЕНИЯ ОБ АНАЛИЗЕ ФИЛОГЕНИИ
Существует распространенное мнение, что реконструирование филогении возможно при сочетании трех взаимодополняющих методов – эмбриологического (см. I.4.2.1), палеонтологического (см. I.4.2.2) и сравнительного (или сравнительно-морфологического).
Сочетание этих трех методов, предложенное во второй половине XIX века Геккелем называют «Геккелевской триадой». Однако из названных трех методов только сравнительный метод (а точнее, филогенетический анализ, или анализ синапоморфий) имеет реальное значение (см. выше, I.4.1). Кроме того, некоторые авторы пытаются привлекать для обоснования филогенетических реконструкций такие, по их мнению, общие закономерности эволюции, как принцип усложнения организации, принцип олигомеризации и другие (см. I.4.2.3, I.4.3.4, I.4.3.5). Однако в действительности каких-либо общих законов эволюции не существует (или, по крайней мере, еще не открыто), и, согласно современным представлениям, эволюционные преобразования могут быть самыми разнообразными. В литературе конца XX века широкое распространение получили нумерические методы реконструкции филогении, не имеющие научного обоснования (см. I.4.2.6). В последнее время большие, но необоснованные надежды возлагают на т.н. «молекулярный метод» реконструкции филогении (см. I.4.2.7).
I.4.2.1. Несостоятельность эмбриологического метода
Эмбриологический метод основывается на «биогенетическом законе», согласно которому каждый организм в процессе своего онтогенеза в сокращенной и измененной форме повторяет свой филогенез. Иными словами, согласно «биогенетическому закону», более древние признаки должны появляться в индивидуальном развитии раньше, чем эволюционно более молодые признаки.
Для обоснования «биогенетического закона» используют специально подобранные примеры, в которых животные из разных таксонов на более ранних стадиях онтогенеза имеют большее сходство между собой, чем те же животные на более поздних стадиях онтогенеза.
Например, во многих учебниках приводится рисунок, показывающий некоторые стадии онтогенеза рыбы, птицы и млекопитающего, при этом на наиболее ранней из изображенных стадий эти животные очень похожи друг на друга (имеют жаберные щели, маленькие конечности и крупный хвост), а на самой поздней из изображенных стадий они резко различаются.
Однако хорошо известны и многочисленные обратные примеры, когда наибольшие различия наблюдаются не на более поздней, а, наоборот, на более ранней стадии онтогенеза.
Например, дифференциация зародышевых слоев (эктодермы, энтодермы и мезодермы) является общим признаком Monostomata, тогда как способы образования этих слоев на более ранних стадиях эмбриогенеза являются признаками подчиненных таксонов [см. главу III: Monostomata (1)]. Даже если обратиться к упомянутому выше хрестоматийному примеру с онтогенезом рыбы, птицы и млекопитающего, то эти же животные на самых ранних стадиях онтогенеза (дробление, гаструла и др.) имеют более резкие внешние различия, чем на стадии жаберных щелей.
Многие исследователи неоднократно пытались использовать «биогенетический закон» для выявления полярности признаков (т.е. для установления, в каком направлении происходила эволюция того или иного признака) и для реконструирования предка. Зачастую это приводило к явно ошибочным гипотезам.
В энтомологии примерами безрезультатных попыток применить биогенетический закон являются существовавшая в XIX веке теория об исходном камподиевидном предке насекомых, основанная на сходстве Campodea и некоторых личинок насекомых (см. главу VI: Diplura: «Статус»); попытки объяснить происхождение крыльев насекомых, исходя из развития крыльев в онтогенезе (см. главу VII: Pterygota: «Классификации» VII); представление о гомологии вентральных придатков стрекоз с церками [см. главу VII: Odonata (10)] и другие. Некоторые авторы считают, что чем раньше в онтогенезе проявляются различия между животными, тем раньше произошла дивергенция этих животных в филогенезе; на основании этого предполагают, в частности, полифилетическое происхождение членистоногих, мотивируя это тем, что у разных членистоногих различаются ранние этапы эмбриогенеза (Anderson 1973). Однако если логически продолжить эту мысль, то мы придем к абсурдному выводу, что разные группы членистоногих и даже разные представители такого явно голофилетического таксона как Collembola (см. главу VI) произошли от разных простейших, поскольку у них описаны разные типы дробления яйца – т.е. процесса перехода от одноклеточного к многоклеточному состоянию. В целом, однако, «биогенетический закон» оказался малопопулярным среди энтомологов, так как глубокая специализация личинок у большинства насекомых наглядно демонстрирует несостоятельность этого закона.
«Биогенетический закон» был сформулирован в XIX веке, когда ничего не было известно о молекулярных носителях наследственности и о механизме их изменения в эволюции. Сейчас мы знаем, что в ходе онтогенеза генотип не меняется, и организм на всех этапах своего онтогенеза строится по программам, закодированным в одном и том же генотипе; в ходе же филогенеза меняется генотип, но при этом изменение генотипа происходит не путем надстройки новых генов, которые бы кодировали вновь добавленные этапы онтогенеза, а путем изменения (мутаций) имевшихся ранее генов. Т.е. современные знания о механизме наследственности никак не проясняют вопрос, каким образом могло бы обеспечиваться повторение филогенеза в онтогенезе. Признаки, сохранившиеся от предка и указывающие на родственные связи данного животного, могут с равным успехом проявиться на любой стадии развития; иногда такие признаки обнаруживаются на ранней стадии, и тогда говорят о проявлении «биогенетического закона»; иногда они, наоборот, выявляются на поздней стадии.
Можно сделать вывод, что эмбриологический метод, как особый метод реконструирования филогении, не обоснован ни теоретическими соображениями, ни эмпирическими данными.
В то же время данные эмбриологии бывают чрезвычайно важны для реконструкции филогении, если они используются не в качестве особого эмбриологического метода, а как материал для филогенетического анализа.
Например, взрослые ракообразные (Eucrustacea) имеют настолько разнообразное строение, что трудно назвать какие-либо общие признаки этого таксона; в то же время у всех Eucrustacea личинки (науплиусы) или поздние эмбрионы имеют явные черты сходства [см. главу IV: Eucrustacea (1)], что позволяет сделать вывод о филогенетической целостности этого таксона. При этом трудно представить себе, что науплиус повторяет строение общего предка ракообразных, поскольку он имеет несегментированное тело и лишь две пары крупных посторальных придатков (рис. 4.7.1E); судя по всему, ракообразные, будучи многосегментными животными, произошли от многосегментного членистоногого предка.
У многих членистоногих в ходе эмбриогенеза сегменты закладываются как гомономные; нередко на определенных стадиях эмбриогенеза сегментация тела выражена отчетливо, тогда как у взрослых животных она местами утрачивается или маскируется – например, фактически несегментированной оказывается голова насекомых и большинства других членистоногих. Благодаря изучению эмбриогенеза удается разобраться в сегментарном составе головы и некоторых других отделов тела, что позволяет выявить общие признаки определенных сегментов и на основании этого делать выводы о филогенетических связях разных групп членистоногих.
I.4.2.2. Ограничения палеонтологического метода
Палеонтологический метод предполагает, что можно непосредственно увидеть результат эволюционного преобразования, если сравнить современных животных с ископаемыми остатками их предков. Действительно, можно найти остатки той особи, которая оставила после себя потомство, давшее начало современным видам. В некоторых случаях сохранность ископаемых остатков и точность установления их возраста достаточны для того, чтобы сделать важные достоверные выводы о строении и времени существования этих животных. Однако в настоящее время нет никаких прямых методов для того, чтобы установить, является ли то или иное ископаемое животное непосредственным предком какого-либо современного животного. Выводы об отношениях предок-потомок можно сделать только с помощью филогенетического анализа. Для этого надо на основе сравнения современных видов реконструировать их общего предка, а затем убедиться в том, что эта реконструкция не противоречит строению данного ископаемого животного. При этом нет никакой гарантии того, что ископаемое животное является непосредственным предком современных, а не членом побочной ветви, сохранившей те черты предка, которые нам удалось реконструировать.
Палеонтологический метод можно понимать в виде такого правила: остатки животных, имеющих плезиоморфии (т.е. предковые черты строения), встречаются преимущественно в более ранних отложениях, чем остатки животных, имеющих апоморфии (т.е. производные черты строения). Однако имеются многочисленные исключения из этого правила, что объясняется прерывистым и случайным характером палеонтологической летописи. Ясно, что если в момент геологической истории А в некоторой филогенетической линии морфологический признак X1 преобразовался в X2, то признак X2 может быть обнаружен у каких-то животных, живших позже момента А, но не может присутствовать ни у каких животных, существовавших до момента А. Однако для распространения во времени признака X1 нет никаких ограничений, так как с появлением признака X2 исходный признак X1 совершенно не обязательно должен исчезать, он может сколь угодно долго сохраняться в филогенетических ветвях, родственных той, где возник признак X2. Поэтому в сформулированном выше правиле фигурирует слово «преимущественно», и если интересующие нас ископаемые животные известны по единичным находкам, это правило не применимо.
Палеонтологическая летопись во многом является случайной по отношению к филогении, но имеет некоторые собственные закономерности (см. главу II.2). Например, мягкотелые животные имеют гораздо меньше шансов сохраниться, чем животные с твердым скелетом, поэтому наличие в древних отложениях животных со скелетом и отсутствие там мягкотелых животных отнюдь не свидетельствует о том, что современные мягкотелые животные возникли в результате утраты скелета. Поскольку наземные животные и растения захоранивались преимущественно в озерных отложениях, наибольшее число их ископаемых остатков относится к озерным и прибрежным видам; но из этого отнюдь не следует, что эти виды древнее всех прочих. Богатые коллекции ископаемых получены из отдельных локальных месторождений, где условия осадконакопления и вся геологическая история способствовали хорошей сохранности ископаемых; но из того факта, что в эти локальные захоронения не попали те или иные виды, отнюдь не следует, что эти виды в то время не существовали. Большое число исключений делает сформулированное выше правило малопригодным.
Примером неудачной интерпретации палеонтологических находок может служить история изучения Palaeodictyoptera и других Protorrhynchota (см. главу VII). Во второй половине XIX – начале XX века общепринятым было мнение, что поскольку эти вымершие насекомые являются одними из самых древних известных крылатых насекомых (они появились в середине карбона и вымерли в перми), эта группа насекомых должна быть и самой примитивной. Палеодиктиоптер называли «синтетической группой», содержащей в себе предков современных отрядов – поденок, стрекоз, прямокрылых, хоботных, перепончатокрылых и др. В соответствии с этим, в частности, в строении Palaeodictyoptera усматривали предковое состояние для стрекоз, и на основании этого наиболее примитивными среди современных стрекоз считали разнокрылых стрекоз (Anisoptera), поскольку они более, чем равнокрылые стрекозы (Zygoptera) похожи на палеодиктиоптер (см. главу VII: Odonatoptera: «Классификация»). Однако, как выяснилось по мере накопления палеонтологического материала, палеодиктиоптеры и другие проторинхоты образуют голофилетическую группу, характеризующуюся уникальной модификацией ротового аппарата и не являющуюся предковой ни для каких современных насекомых; соответственно, потеряли свое значение и те теории о филогении отдельных групп крылатых насекомых, которые были основаны на представлении о происхождении их от палеодиктиоптер. Причиной ошибки, по-видимому, было то, что крупные хорошо летавшие палеодиктиоптеры захоранивались в озерных отложениях карбона, тогда как по-настоящему примитивные крылатые насекомые вели иной образ жизни и в эти отложения не попали.
Палеонтологический метод как особый метод реконструирования филогении не дает желаемых результатов. В то же время данные палеонтологии имеют огромное значение для реконструирования филогении. Во многих случаях ископаемые животные имеют такие признаки и сочетания признаков, которые не встречаются у современных видов. Поэтому сравнительный анализ ископаемых дает результаты, которые нельзя было бы получить, изучая лишь современные формы. Однако при этом используется не особый палеонтологический метод, а филогенетический анализ, применяемый к палеонтологическим объектам.
I.4.2.3. Необязательность усложнения строения
Бытует мнение, что эволюционные преобразования происходят в направлении усложнения строения организмов. Такой «закон» обязательного или преимущественного усложнения организации в ходе эволюции мог бы быть объясним, если исходить из представлений Ж.Б. Ламарка, актуальных в конце XVIII – начале XIX века.
Согласно этой старинной теории, на организм извне воздействуют флюиды, на что организм каким-то образом адекватно реагирует так, что в нем образуются каналы для проведения этих флюидов и для приспособления их к нуждам организма (т.е. сосуды, нервы и прочие интегрирующие системы); это усложнение строения каким-то образом передается по наследству. В результате при адаптации к новым условиям среды, помимо существующих уже структур, добавляются новые, т.е. новая адаптация ведет к усложнению строения, что и приводит к закономерному преобразованию более простых организмов в более сложные. Однако теория флюидов была опровергнута еще во времена Ламарка, а его представления об адекватной реакции организма на воздействия внешней среды и о наследовании приобретенных признаков в дальнейшем не нашли подтверждения.
Не вызывает сомнений, что первые живые организмы имели простое строение; что более сложно устроенные эукариоты произошли из более просто устроенных прокариот; что многоклеточные животные произошли из одноклеточных, и что предками таких сложно устроенных многоклеточных, как членистоногие или позвоночные, были многоклеточные с более простым строением. Поскольку исходно организмов со сложным строением не было, они появились именно в результате усложнения. Однако из этого никак не следует, что существует закон усложнения организации: эволюция идет с равным успехом как в сторону усложнения, так и в сторону упрощения. Когда мы рассматриваем филогенетические отношения каких-либо животных, из которых одни имеют более сложное строение, а другие более простое, мы можем с равным успехом предположить и происхождение более сложного из более простого, и наоборот, более простого из более сложного. Поэтому сравнение организмов по их сложности само по себе не позволяет реконструировать филогению.
Рассматриваемое ниже представление о «законе» олигомеризации гомологичных органов является противоположностью «закона» об усложнении организации.
I.4.2.4. Необязательность олигомеризации
Многие современные филогенетические теории, в том числе в энтомологии, основаны на предположении, что в ходе эволюции происходит олигомеризация (т.е. уменьшение в числе) тех или иных органов, изначально присутствовавших в большом числе. Например, предполагается, что у членистоногих исходно было большое число одинаковых пар ног, а в дальнейшем число пар ног сокращалось (см. Euarthropoda: «Классификации»); у крылатых насекомых исходно было две пары одинаковых крыльев, а в дальнейшем остается одна пара (см. главу VII) и т.п. В большинстве своем эти теории являются вполне обоснованными, так как аргументированы с позиций филогенетического анализа (см. ниже).
Однако следует предостеречь от абсолютизации идеи об олигомеризации гомологичных органов. Когда рассматривают филогению отдельной группы животных, стараются из всех возможных объяснений существующего строения животных выбирать наиболее простое. Если у одного животного имеется некий орган, а у другого животного этого органа нет, то проще предположить, что произошла редукция этого органа у второго животного, нежели предложить гипотезу, объясняющую появление этого органа у первого. Так, идя по пути наиболее легкого объяснения, некоторые авторы пытаются все эволюционные преобразования свести к редукциям.
Например, в работах Кукаловой-Пек (Kukalova-Peck 1983, 1985, 1987, 1991) предложена теория филогении крылатых насекомых (Pterygota), в которой гипотетическая предковая форма несет на себе чуть ли не все детали строения, найденные на разных сегментах разных насекомых, причем в полимеризованном виде. На каждом его сегменте, помимо пары конечностей, имеются и гомологи глаз, и гомологи крыльев; каждая конечность имеет максимальное число члеников и коготков, известное для каких-либо конечностей какого-либо насекомого; каждый членик несет стилус, каждый стилус членистый; у крыла каждой жилке соответствует свой набор аксиллярных склеритов, и т.п. Рассуждая таким образом, автор освобождает себя от необходимости объяснить появление в эволюции новых органов: в данном случае вопросы о возникновении деталей строения, характерных для Pterygota, предлагается решать кому-то другому – тому, кто реконструирует более ранние этапы филогении членистоногих.
Однако очевидно, что поскольку те или иные органы у животных существуют, их возникновение так или иначе придется объяснить.
В некоторых случаях филогенетическая гипотеза, основанная на представлении об олигомеризации гомологичных органов, оказывается несостоятельной.
В прошлом многие авторы рассматривали многоножек (Myriapoda) с большим числом сегментов туловища как примитивные формы. В частности считалось, что Epimorpha являются наиболее примитивными среди Chilopoda, так как имеют большое варьирующее число сегментов (см. главу V: Chilopoda: «Классификации» I). Однако с позиций филогенетического анализа эта версия выглядит невероятной, и приходится принять гипотезу, согласно которой увеличение числа сегментов у Chilopoda произошло вторично (см. там же, II).
Подобным образом, согласно существующим ныне представлениям о филогении Diplopoda, большое неопределенное число сегментов, свойственное кивсякам (в составе Helminthomorpha), является вторичным, тогда как исходно диплоподы имеют сравнительно небольшое постоянное число сегментов (см. главу V).
Таким образом, сравнение животных по их олигомерности и полимерности само по себе недостаточно, чтобы делать вывод о направлении эволюции.
Нередко предковый вид какого-либо таксона представляют как нечто неспециализированное, совмещающее в себе черты сразу всех членов таксона. С этим представлением связан иногда употребляемый термин «генерализованный» (то есть «обобщенный») признак или организм. На самом же деле любой вид животных имеет свои особенности, свою специализацию, и эволюционирует под действием естественного отбора в частных условиях среды, а не под влиянием стремления породить в будущем большой и разнообразный таксон. В ходе эволюции происходит не только специализация, но и деспециализация, и изменение специализации.
В качестве одного из многочисленных примеров исчезновения и изменения исходной специализации можно привести эволюцию личинок поденок (Ephemeroptera): убедительно показано, что личинки поденок исходно имели специализацию к активному плаванию [см. главу VII: Ephemeroptera (1e)], которая в разных группах поденок исчезала или заменялась на специализацию к иному образу жизни; иногда такие личинки, утратившие плавательную специализацию, кажутся «генерализованными», так как напоминают многих насекомых сразу.
Не вызывают также сомнений теории, согласно которым эволюционный путь, приведший к пчелам с их социальным образом жизни и питанием пыльцой растений, проходил через специализацию к паразитизму на насекомых (свойственную примитивным стебельчатобрюхим перепончатокрылым); путь, приведший к четвероногим и человеку, проходил через специализацию к плаванию (свойственную рыбообразным), и многие другие.
По всей видимости, неизвестные нам предковые виды, такие как общий предок насекомых, общий предок крылатых насекомых, общий предок насекомых с полным превращением и другие, были вполне специализированными насекомыми, а такие фундаментальные признаки крупных таксонов, как крылья или полное превращение, исходно возникли как частные адаптации к конкретным условиям среды, а не как необъяснимое стремление породить биологическое разнообразие.
I.4.2.6. Причины несостоятельности нумерических и матричных методов
Четкая формулировка принципов филогенетического анализа вызвала у многих исследователей соблазн формализовать процесс реконструкции филогении и использовать для этой цели компьютер. В настоящее время составлены компьютерные программы для построения филогенетических древес на основе заранее выявленных апоморфий. Однако реконструирование филогении формализованными методами наталкивается на непреодолимые трудности и вряд ли является целесообразным. Поскольку увлечение этими методами сейчас широко распространено, здесь следует разобраться в их принципиальных недостатках.
Для компьютерного построения филогенетического древа сначала составляют матрицу таксон/признак (что само по себе некорректно – см. ниже, I.4.2.6.2), где для каждого включенного в анализ таксона указывают состояние признака и отмечают, является ли это состояние признака апоморфией или плезиоморфией. Затем на основе этой матрицы строится филогенетическое древо; программа построения древа базируется на том же принципе, что и настоящий филогенетический анализ (см. выше, I.4.1) – т.е. на предположении, что все одинаковые апоморфные состояния признаков являются синапоморфиями, т.е. унаследованы от общего предка. Если бы все этим и ограничивалось, использование компьютерной программы было бы явно излишним, так как все эти процедуры легче сделать вручную, чем на компьютере. Однако при построении древа на основе матрицы таксон/признак зачастую обнаруживаются нерешаемые комбинации признаков:
|
Допустим
что: |
Виды: |
Признаки: |
|
|
x |
y |
||
|
A |
x1 |
y2 |
|
|
B |
x2 |
y1 |
|
|
C |
x2 |
y2 |
|
Для такой матрицы ни одного филогенетического древа построить невозможно, так как в любом случае получится, что какой-то признак (либо x2, либо y2) возник независимо два раза. Здесь можно предположить, что в каком-то случае неправильно определена полярность признаков. Если изменить полярность какого-либо признака (например, x) на противоположную, то на основе этой же матрицы можно построить непротиворечивое филогенетическое древо:
|
Допустим, что |
|
Можно использовать программу, в которой полярность не задается исходно, а выявляется в ходе построения филогенетического древа: правильной будет считаться та полярность признаков, которая позволяет построить непротиворечивое древо (как, например, в предыдущем примере). Однако в матрице может оказаться такое сочетание признаков, при котором непротиворечивое филогенетическое древо не может быть построено ни при каком допущении о полярности признаков:
| Допустим
что: один из признаков x1 или x2 – апоморфия, другой – плезиоморфия; один из признаков y1 или y2 – апоморфия, другой – плезиоморфия. |
Виды: | Признаки: | |
| x | y | ||
| A | x1 | y2 | |
| B | x2 | y1 | |
| C | x2 | y2 | |
| D | x1 | y1 | |
Теоретически таких нерешаемых сочетаний признаков быть не должно, но на практике они встречаются очень часто. Причины существования таких сочетаний могут быть различными. 1) В результате конвергенции, при адаптации к одинаковым условиям, могут независимо возникать сходные признаки, которые кажутся идентичными, но имеют разную генетическую основу; такие случаи вызывают затруднения не только при компьютерном, но и при любом другом методе реконструирования филогении. 2) Идентичные признаки, имеющие одну и ту же генетическую основу, могут независимо проявляться в родственных группах (см. выше, I.4.1.4). 3) То, что кажется синапоморфией, может быть на самом деле искусственным признаком – т.е. несколькими совершенно разными, но одинаково сформулированными признаками. Если первые две причины создают трудности для любого филогенетического анализа, то последняя из этих причин характерна именно для обработки данных формализованными компьютерными методами: исследователь может отличить явно искусственный признак от реального, но компьютер, имеющий дело не с признаками, а только с их формулировками, этого сделать не в состоянии. При обнаружении нерешаемого сочетания признаков необходимо переисследовать эти признаки у изучаемых животных для того, чтобы попытаться выявить допущенную ошибку. Однако компьютерные программы рассчитаны не на переисследование животных, а на попытку решить проблему чисто компьютерными способами.
Чтобы обойти возникшее препятствие, вводится допущение, что эволюция может быть обратимой и/или повторимой. Если не добавлять к этому никаких дополнительных условий, филогенетический анализ теряет всякий смысл, и компьютер строит множество возможных древес. Для того, чтобы выбрать из множества древес единственное, используют принцип, называемый принципом экономности, или парсимонии (parsimonia). Он выражается в том, что выбирается такое древо, в котором нарушение принципа необратимости и/или неповторимости происходит наименьшее число раз.
I.4.2.6.1. Несостоятельность принципа экономности. Принцип экономности не имеет отношения ни к каким биологическим теориям. Обычно, ссылаясь на принцип экономности при реконструкции филогении, не уточняют, экономность чего именно имеется в виду – то ли это экономность природы, то ли экономность в действиях исследователя.
В первом случае принцип экономности находится в прямом противоречии с современной наукой. Выбранное на его основе наиболее экономное филогенетическое древо ни на каком основании нельзя считать правильным, поскольку в природе неизвестен механизм, который заставлял бы животных избирать наиболее экономный (или вообще оптимальный в каком бы то ни было отношении) путь эволюции. Филогения всякой группы животных является уникальной. Каждый эволюционный процесс произошел один раз, и поэтому вообще не может существовать какого-либо механизма оптимизации эволюционных процессов. Можно было бы выдвинуть какую-нибудь теорию экономной эволюции, альтернативную дарвиновской; в этом случае использование принципа экономности для реконструкции филогении конкретной группы животных опиралось бы на выдвинутую теорию и, таким образом, находилось бы в рамках научного познания. Однако те авторы, которые ссылаются на принцип экономности при реконструкции филогении, одновременно с этим ссылаются на теорию Дарвина как на единственную возможную. Такой подход прямо противоречит принципу научного познания.
Во втором случае представление о том, что исследователь должен действовать экономно (то есть не придумывать лишних сущностей там, где можно без них обойтись) никак не вяжется с теми действиями, которые совершают исследователи при анализе филогении матрично-нумерическими методами. Для вычисления «экономности» написано огромное количество весьма сложных программ, пользование которыми уже само по себе требует определенных знаний и умений.
В качестве единственного обоснования использования принципа экономности в филогенетическом анализе ссылаются на работы философа К. Поппера. Этот автор, не будучи биологом, не имел представления о проблемах анализа филогении, но в его публикациях содержатся рассуждения о методах познания в физике. Объясняя способ познания закономерно повторяющихся физических явлений, Поппер ссылался на принцип опровергаемости научных теорий (почему-то названный им принципом «фальсифицируемости», хотя в общепринятой лексике это слово имеет противоположное значение). Согласно этому принципу, универсальная научная теория (т.е. теория, объясняющая бесконечное множество частных фактов) в принципе не может быть доказана, поскольку для этого понадобилось бы рассмотреть бесконечно много фактов; но она может быть опровергнута: для этого достаточно найти один факт, который ей противоречит. Поэтому научная гипотеза должна быть опровергаемой, т.е. должна быть сформулирована так, чтобы в принципе можно было бы предположить существование каких-то фактов, которые ей противоречат. До тех пор, пока таких фактов обнаружить не удается, гипотеза считается правильной (точнее, не опровергнутой). Для объяснения одного и того же явления можно выдвинуть много различных гипотез, для опровержения каждой из которых понадобится затратить некоторое время. Чтобы упорядочить этот процесс, в качестве рабочей гипотезы целесообразно выбрать наиболее экономную (т.е. наиболее простую) из всех возможных, а в случае ее опровержения – выбрать наиболее экономную из оставшихся гипотез. Здесь принцип экономности не является критерием истинности той или иной гипотезы, а лишь используется для выбора последовательности, в которой должна проверяться истинность различных гипотез. Когда же говорят о принципе экономности при анализе филогении, подразумевают нечто прямо противоположное: экономность используют как единственный и окончательный критерий для выбора «истинной» филогенетической гипотезы среди множества гипотез, ни одна из которых в дальнейшем проверяться не будет (Песенко 1989).
I.4.2.6.2. Некорректность представления данных в виде матрицы. Для использования нумерических методов вначале составляют прямоугольную двухмерную матрицу таксон/признак (см. I.4.2.6). На первый взгляд такое представление данных кажется разумным; иногда исследователю действительно бывает полезно составить для себя такую таблицу признаков, которая воспринимается легче, чем набор словесных описаний таксонов.
Однако нельзя сводить всю информацию о распределении признаков к матрице.
Во-первых, при составлении матрицы приходится придумывать новый искусственный набор признаков. Как показал опыт биологической систематики со времен Линнея, и как явствует из современной теории эволюции, разнообразие живых организмов укладывается не в матричную, а в иерархическую форму. Линней совершенно справедливо писал в «Философии ботаники»: «то что в одном роде важно для установления рода, в другом – вообще не имеет значения». Систематики формулируют каждый признак таким образом, что при данной формулировке он оказывается стабильным в пределах определенного таксона и позволяет надежно отличить этот таксон от одного или нескольких ближайших к нему таксонов. Однако это не означает, что та же формулировка признака позволит охарактеризовать другие таксоны.
Например, таксон Raptoriae (богомолы) четко характеризуется определенной хватательной специализацией передних ног [см. главу VIII: Raptoriae (1)]; однако у других насекомых передние ноги бывают самыми разными – от неспециализированных до таких же хватательных, как у богомолов, так что этот признак невозможно занести в матрицу для всех насекомых.
Чтобы свести все признаки в строгую двухмерную матрицу, приходится применять искусственные приемы – выбрасывать неудобные признаки и изменять формулировки признаков, отказываясь от тех, что уже оправдали себя при построении иерархической системы. В результате этого, когда на следующем этапе анализа из матрицы получают иерархическую систему (в виде ветвящегося филогенетического древа), она оказывается основанной на меньшем числе фактов, чем было известно исходно.
Зачастую для составления матрицы сложный признак заменяют набором простых признаков, полагая, что такое действие является анализом. В действительности такое разложение признака на составляющие является не естественно-научным анализом, а лишь чисто логическим приемом, в результате которого информация не добавляется, а, наоборот, исчезает. Например, если некая часть тела животного имеет характерную сложную форму, мы понимаем, что такая форма не может повторяться случайно, и поэтому успешно используем этот признак в филогенетическом анализе. Однако внести такую информацию в матрицу невозможно: если мы обозначим данную сложную форму одним условным символом и запишем его в клетки матрицы, относящиеся к видам, обладающим этой формой, то мы не сможем заполнить соответствующие клетки для других видов. Чтобы заполнить все клетки матрицы, приходится заменить обозначение уникальной формы на набор эпитетов, каждый из которых имеет свой антоним: например, «длинный» (антоним – «короткий»), «расширен дистально» (антонимы – «не расширен дистально» и «сужен дистально»), «с выступом» (антоним – «без выступа») и т.п. В этом случае все клетки матрицы оказываются заполненными, но при этом признаки, которыми они заполнены, утратили свою уникальность и поэтому стали непригодными для филогенетического анализа.
Во-вторых, при составлении матрицы приходится придумывать новый искусственный набор таксонов. Таксоны, заносимые в матрицу, называют «операциональными единицами». Операциональные единицы могут быть таксонами одного ранга или иметь разные ранги в исходной формальной классификации (т.е. среди них могут быть виды, роды, семейства и др.); в обоих случаях далее все они анализируются как бы имеющие один и тот же условный ранг. Выбор таксонов, которые уравниваются в ранге, является произвольным (как и в традиционной ранговой классификации – см. I.5.4.2).
Поскольку от выбора операциональных единиц зависит формулировка признаков (см. выше), матрица оказывается в значительной мере произвольной и искусственной.
Зачастую матрицу составляют для того, чтобы обрабатывать ее математическими методами, основанными на «принципе экономности». Такое действие явно абсурдно, поскольку единственным оправданным пониманием «принципа экономности» является экономность в действиях исследователя (см. I.4.2.6.1), а составление матрицы вместо дендрограммы с целью получить дендрограмму, так же как и применение громоздких математических вычислений, никак нельзя назвать экономными действиями.
Составление матрицы таксон/признак вполне оправдано в тех случаях, когда в нее заносят признаки, имеющие мозаичное распределение по таксонам. Мозаичное распределение признака получается, если генная программа, кодирующая признак, имеется во всем множестве этих таксонов, но реализуется только у части в них. Признаки, распределенные таким образом, не могут быть использованы в филогенетическом анализе.
I.4.2.6.3. Фундаментальные недостатки нумерических методов в систематике. Использование нумерических (в частности компьютерных) способов реконструирования филогении лишено смысла по следующим причинам.
(А) В рассмотренных выше методах, как и во многих других, все вычисления делаются на основе произвольно составленных «удобных» математических формул; такой род занятий нельзя считать научным. В науке применение математики выражается в том, что научная теория записывается в виде математической формулы, и именно эта формула используется для вычисления в случаях, предусмотренных данной теорией. В свою очередь, теория, отраженная в формуле, является предметом научных дискуссий, поскольку сформулирована в такой форме, что в принципе допускает возможность ее опровержения.
(Б) В большинстве применяемых сейчас математических методов реконструирования филогении используется такая величина, как число признаков – число общих или различающихся признаков нескольких таксонов, число апоморфий или плезиоморфий и др.
Филогенетическое древо, построенное по этим методам, зависит от числа синапоморфий у тех или иных таксонов. Здесь обратим внимание на то, что в отличие от нумерического, в настоящем филогенетическом анализе (см. I.4.1) число синапоморфий никак не влияет на результат: там филогенетическая ветвь считается существующей, если ее члены имеют хотя бы одну синапоморфию; большее число синапоморфий для этих же групп придает больше уверенности в правильности реконструированного древа, но не влияет на само древо. Используемое в нумерических методах число признаков является понятием, бессмысленным с биологической точки зрения, так как один и тот же признак можно сформулировать различным образом, кратко или длинно.
Например, признак таксона Entognatha [см. главу VI: Entognatha (1)] можно записать в виде одного слова: «скрыточелюстность». Этот же признак можно записать значительно длиннее (см. там же). Длинную запись можно сделать в виде одного сложносочиненного предложения, и тогда это будет считаться одним признаком. Можно эту же запись сделать в виде многих предложений, и пронумеровать каждое в отдельности, тогда все они будут считаться разными признаками. Точно так же признак таксона Pleomerentoma, касающийся строения придатков брюшка (см. главу VI: Hexapoda: «Классификации» III), можно записать в виде одного или нескольких предложений, считая их одним или несколькими признаками. Если предложений (т.е. подсчитываемых «признаков») в характеристике Entognatha окажется больше, чем в характеристике Pleomerentoma, получится следующая филогенетическая схема:

Если наоборот, получится совсем другой вывод о филогении:

Даже если мы возьмем совершенно простые предметы, например квадрат и круг, мы не сможем однозначно ответить на вопрос, сколько между ними различий, т.е. сколькими особыми признаками обладает каждый из них.

Можно сказать, что здесь различие одно – первый предмет квадратный, а второй круглый. Можно сказать, что различий 4 – в первом случае четыре угла, во втором ни одного; можно с тем же успехом посчитать и углы, и стороны, кривизну сторон и т.п. На самом деле каждый геометрический предмет включает бесконечное число точек и поэтому описывается уравнением, имеющим бесконечное число значений.
Строение организма определяется генами; число генов многоклеточного организма очень велико, но не бесконечно. Мы же учитываем не гены, а признаки; в отличие от генов, число признаков не велико, а бесконечно.
В нумерических методах с числами признаков производятся арифметические действия – деление, вычитание и т.п. В действительности, числа признаков во всех случаях равны бесконечности, и совершать над ними арифметические действия может лишь человек, не понимающий, что он делает.
Все сказанное здесь о подсчете признаков относится не только к нумерическим методам реконструирования филогении, но и к любым другим нумерическим методам в систематике. В 60-е годы существовало такое течение, как фенетическая, или нумерическая систематика, при котором отрицалось значение филогении для систематики и предлагалось основывать классификацию на «объективных» вычислениях. Современные методы нумерической кладистики во многом заимствованы у фенетики (Майр 1971, Песенко 1989–1991).
(В) Независимо от того, применяются или нет неадекватные математические методы (см. А) и используется или нет искусственный подсчет признаков (см. Б), применение компьютера для реконструирования филогении не может принести желаемого результата.
Все преимущество компьютера состоит в том, что он способен быстро выполнять длинную цепь логических операций; если же требуется выполнить подряд лишь несколько логических операций, то это легче сделать в уме. Если бы можно было разбить процесс реконструирования филогении на два последовательных этапа – на первом этапе описание (то есть формализация) признаков, а на втором этапе реконструирование филогении на основе этих формализованных признаков, то второй этап можно было бы выполнять с помощью компьютера. Однако сформулировать признаки можно только имея представление о филогении изучаемой группы, поскольку всякое описание животного и всякая формулировка признака уже содержит представление автора о природе этого животного или признака (см. выше, I.4.1.3). Таким образом, вся работа по реконструированию филогении должна делаться одновременно, без разбиения на этапы. Это значит, что после короткой последовательности логических операций (настолько короткой, что ее легко сделать в уме) необходимо возвращаться к переисследованию строения животных с целью уточнить или исправить формулировки признаков.
Например, если мы получили два противоречивых древа, показанных выше, бесполезно продолжать логические операции с имеющимися в нашем распоряжении формулировками признаков Entognatha и Pleomerentoma. Вместо этого нужно вскрыть головы различных представителей Entognatha, сравнить их с другими насекомыми и, учитывая возникшие сомнение в единстве происхождения Entognatha, заново описать строение их ротовых аппаратов. То же нужно проделать и с придатками брюшка различных насекомых, с целью критически переосмыслить признаки Pleomerentoma.
I.4.2.7. Подмена молекулярного метода анализа филогении
Стремительное увеличение знаний о строении и функции ДНК позволяет надеяться, что эти знания могут быть использованы для понимания филогении. В то же время современная научная литература переполнена публикациями по так называемой «молекулярной филогении», не имеющими научного значения.
Часто употребляемое словосочетание «молекулярная филогения» нельзя понимать буквально, потому что филогения (т.е. образование родственных групп) может быть только у целостных животных, имеющих родство в буквальном смысле слова, но не у их молекул или каких-либо других частей. Под «молекулярной филогенией» подразумевают молекулярные методы реконструирования филогении, точнее, методы реконструирования филогении, основанные на анализе ДНК. В настоящее время об этих методах пишут так много, что некоторые люди даже думают, что можно усовершенствовать анализ ДНК до такой степени, чтобы автоматически получать филогенетическое древо. Однако филогенетическое древо не записано в ДНК, так что при любом совершенстве методов филогенетическое древо останется лишь гипотезой, которую нельзя окончательно доказать, но которую можно бесконечно приближать к действительности, дополняя одни данные другими.
Существует мнение, что молекулярные методы анализа филогении способны уже сейчас заменить и вытеснить анализ филогении, основанный на фенотипических признаках. Такое мнение может возникнуть в результате следующих рассуждений.
При реконструировании филогении традиционным сравнительно-морфологическим методом мы используем только наследуемые морфологические признаки, и только для того, чтобы, анализируя наследование этих признаков, выявить родственные связи организмов. При этом все ясно понимают, что морфологические признаки как таковые не наследуются: когда животное умирает, его тело разрушается или поедается представителями других видов, так что никакие его морфологические части не передаются его потомкам. Помимо одной яйцеклетки (которая не имеет строения взрослого животного), потомок получает в наследство только информацию, записанную в молекулах ДНК. Таким образом, при реконструкции филогении мы анализируем морфологические признаки только для того, чтобы по ним судить о последовательности нуклеотидов в ДНК. Знание о наличии у разных животных одинаковых морфологических признаков (среди которых имеются интересующие нас синапоморфии) нужны нам только для того, чтобы на их основе сделать косвенный вывод о существовании у этих животных одинаковых наследуемых генов, кодирующих эти признаки. В свою очередь, наличие одинаковых генов интересует нас только потому, что на его основе мы можем сделать косвенный вывод об общем происхождении тех животных, которые обладают одинаковыми генами. Таким образом, при анализе филогении с помощью морфологических признаков мы два раза делаем косвенный вывод: (1) предполагая, что в данном случае одинаковые признаки кодируются одинаковыми генами и (2) предполагая, что в данном случае одинаковые гены унаследованы от общего предка. То есть при таком анализе мы имеем два источника возможных ошибок. Если вместо того, чтобы выявлять морфологическое сходство, чтобы по нему косвенно судить о генном сходстве, мы сразу выявим генное сходство путем непосредственного секвенирования ДНК, мы устраним один из этих двух источников ошибок. Второй источник ошибок при этом остается, поскольку ни в морфологии, ни в генах родословная не записана, и в любом случае ее можно выявить только косвенным путем, т.е. выдвигая гипотезы, которые могут оказаться ошибочными. Таким образом, может быть создан особый молекулярный (генотипический) метод реконструкции филогении, альтернативный морфологическому (фенотипическому) методу: при молекулярном методе вместо анализа апоморфий на морфологическом уровне те же апоморфии анализируются на генном уровне. Такой метод был бы принципиально более точным и надежным, чем применяемый ныне морфологический метод (хотя молекулярный метод, как и любой другой, не позволит безошибочно реконструировать филогению с заданной точностью). Если бы молекулярный метод существовал, можно было бы рассуждать о том, в каких случаях какой из двух методов лучше – более дорогой, но более надежный молекулярный, или более доступный, но менее надежный морфологический.
Однако в настоящее время такого молекулярного метода еще нет, как нет и научных знаний, на которых он должен быть основан. Современная наука знает только о том, каким образом гены кодируют белки, и имеет некоторое представление о механизмах взаимодействия генов. О кодировании генами морфологических признаков мы знаем только то, что они их как-то кодируют, но не имеем представления о том, как именно. Ничего неизвестно о том, какие гены кодируют те морфологические признаки, которые используются нами при филогенетическом анализе.
В настоящее время широко практикуется подмена настоящего молекулярного метода его суррогатом: вместо анализа тех генов, которые кодируют интересующие нас апоморфии, делают анализ тех генов, которые для этого доступны – таких как ген рибосомальной РНК, гены определенных энзимов или митохондриальные гены. В этих случаях исследование молекул не представляет собою особого молекулярного метода анализа филогении, который можно было бы противопоставить морфологическому методу. Действительно, если нас интересуют крупные детали строения животного, мы можем просто рассмотреть их своими глазами, а если нас интересует то, что невооруженным глазом не видно, мы используем те или иные технические средства; но применение этих средств не является особым методом филогенетического анализа. Если человек плохо видит, он надевает очки; если его интересуют мелкие детали строения, он использует микроскоп. Если светового микроскопа недостаточно, используют электронный; однако в электронный микроскоп наш глаз ничего увидеть не может, так что мы судим о строении объекта косвенно, на основании реконструкции изображения, создаваемой этим прибором. Для того, чтобы рассмотреть признаки, по которым различаются рибосомы, разрешения электронного микроскопа недостаточно, и об этих признаках можно судить косвенно, на основании секвенса ядерной ДНК, кодирующей рибосомальную РНК. Как и в случае очков, микроскопа или электронного микроскопа, секвенирование ДНК в данном случае является лишь техническим методом изучения морфологии данного объекта. Когда под видом особого «молекулярного метода» анализа филогении вместо традиционных морфологических признаков рассматривают 18S рибосомальную ДНК, это является не заменой морфологического метода молекулярным, а лишь заменой традиционных признаков признаками строения 18S субъединицы рибосомы.
Изучение рибосомальной РНК, энзимов, митохондриальной ДНК или других структур, доступных для изучения молекулярными методами, может быть полезным. Эти данные можно с успехом использовать в филогенетическом анализе, если правильно понимать их значение. Например, если, помимо ранее известного внешнего строения животного, мы получаем знания об особенностях строения его рибосом, мы можем попробовать найти апоморфии, полезные для филогенетического анализа, не только среди признаков внешнего строения, но и среди деталей строения рибосомы. Если в изученном рибосомальном гене такие апоморфии удастся найти, их можно использовать в реконструкции филогенетического древа, в котором отдельные филогенетические ветви доказываются теми или иными признаками внешней морфологии и/или последовательности нуклеотидов в рибосомальной ДНК. Как и любой другой признак, молекулярный признак может быть использован при реконструкции филогении в дополнение к другим используемым для этого признакам.
Вместо этого во многих публикациях фигурируют дендрограммы, выдаваемые за «филогенетические деревья», построенные исключительно на основе данных о секвенсе нескольких известных генов для отдельных видов, при полном и преднамеренном игнорировании всех прочих известных фактов.
По аналогии с известным в систематике представлением о консервативности определенных признаков (см. I.2.4), некоторые авторы обосновывают выбор гена для филогенетического анализа тем, что этот ген якобы обладает большей консервативностью, чем другие. Подобные утверждения не вытекают из эмпирических данных, а основаны лишь на ошибочном представлении о том, что консервативным должен быть тот ген или признак, который не влияет на взаимодействие организма и внешней среды и потому не подвержен естественному отбору. Из опыта построения классификации и анализа филогении по фенотипическим признакам мы знаем, что это не так.
Может создаться впечатление, что если применить к последовательностям нуклеотидов процедуру формального филогенетического анализа, то результаты такого анализа получатся однозначными и объективными. В то время как для математической обработки фенотипических признаков их приходится сначала искусственно формулировать (в результате чего признаки лишаются объективности, и применение математики становится бессмысленным), последовательность нуклеотидов можно записать объективно и однозначно. Однако в действительности мутации, благодаря которым возникло различие нуклеотидных последовательностей, происходили у предков изучаемых животных на фоне сложной генной регуляции, так что замены тех или иных нуклеотидов в ходе этих мутаций не были равновероятными. Из сравнительной морфологии хорошо известно, что определенный признак в определенном таксоне может быть очень устойчивыми и не нарушается никакими мутациями, тогда как подобный признак в другом таксоне может быть очень изменчивым (см. I.2.4). Именно отдельные устойчивые признаки конкретных таксонов (а не любые признаки) традиционно являются основой для реконструкции филогении и построения классификации [«то, что в одном роде важно для установления рода, в другом – вообще не имеет значения» (Линней 1751)]. Для выявления молекулярных механизмов, обеспечивающих устойчивость отдельных признаков, вероятно, недостаточно знания последовательности нуклеотидов, а необходимо разобраться в сложных молекулярных взаимодействиях и выдвинуть гипотезы о том, какими были эти взаимодействия у предков. Так что попытки некоторых авторов вычислить филогению простыми математическими средствами, основываясь лишь на сырых результатах секвенирования ДНК, являются преждевременными.
О том, насколько неправдоподобна современная т.н. «молекулярная филогения», можно судить по такому сопоставлению. В 18S рибосомальной ДНК эукариот содержится 1900 нуклеотидов. Поскольку имеется всего 4 типа нуклеотидов, то каждый из них несет 2 бита информации, так что вся 18S рибосомальная ДНК содержит 3800 бит информации. Сравним это с информацией, содержащейся в тексте. Если в алфавите имеется более 32 букв, то каждая из них несет более 5 бит информации; так что текст, содержащий 3800 бит информации, должен состоять не более чем из 760 букв. В книге среднего формата такой текст займет около 12 строчек. Даже если бы кто-нибудь попытался умышленно записать в 18S рибосомальной ДНК все сведения о филогенетическом положении обладателя этой рибосомы, ему вряд ли удалось бы вместить в нее такое количество зашифрованной информации (не говоря уже о том, что рибосома, помимо сомнительной функции исторического архива, должна еще и белок синтезировать). Тем не менее, анализируя филогению по одним только 18S рибосомальным ДНК, некоторые авторы умудрились у одного и того же животного найти информацию о его филогенетическом положении на всех уровнях – от исходных дивергенций живых организмов до родственных связей с ближайшими видами и популяциями, и написали об этом несравненно больше двенадцати строчек текста.
В настоящее время, несмотря на бурное развитие молекулярных методик и усиленные попытки использовать молекулярные данные для реконструкции филогении, вклад этих исследований в реконструкцию филогении невелик. В частности, в обильной литературе по «молекулярной филогении» насекомых мне не удалось найти ни одного факта, который можно было бы использовать в данной книге.