КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ
Глава I из книги «Систематика насекомых и принципы кладоэндезиса»

![]()
|
КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ Глава I из книги «Систематика насекомых и принципы кладоэндезиса» |
|||||||
|
|
|
fg | |||||
![]() I.1. Что такое
кладоэндезис
I.1. Что такое
кладоэндезис
 I.2. Понятия и
термины, связанные с биологической
эволюцией
I.2. Понятия и
термины, связанные с биологической
эволюцией
![]() I.3. Биологический вид
I.3. Биологический вид
![]() I.4.
Реконструирование
филогении
I.4.
Реконструирование
филогении
 I.5.
Принципы классификации надвидовых
таксонов
I.5.
Принципы классификации надвидовых
таксонов
I.5.1. Понятия моно-, поли-, пара- и голофилии
I.5.1.1. История вопроса
I.5.1.2. Определения
I.5.1.3. Разъяснение понятий голо-, пара- и полифилии
I.5.2. Разные подходы к классификации
I.5.2.1. Кладизм, или филогенетическая систематика
I.5.2.2. Традиционализм, или эволюционная систематика
I.5.2.3. Градизм
I.5.3. Принципы классификации с точки зрения кладоэндезиса
I.5.3.1. Польза и вред парафилетического (предкового) таксона
I.5.3.2. Плезиоморфоны
I.5.3.3. Стремление классификации к кладистическому идеалу
I.5.3.4. Причины разногласий между кладистами и традиционалистами
I.5.4. Относительные и абсолютные ранги таксонов
I.5.4.1. Строгая иерархичность классификации
I.5.4.2. Абсолютные ранги
I.5.4.3. Неравноценность абсолютных рангов
I.5.4.4. Проблема рода
![]() I.6.
Принципы номенклатуры таксонов
I.6.
Принципы номенклатуры таксонов
I.5. Принципы классификации надвидовых таксонов
Подавляющее большинство современных исследователей сходится на том, что классификация должна отражать филогению, поскольку только филогенетические связи между видами животных могут быть основой для построения их естественной классификации. Принимая этот тезис, следует, однако ясно понимать, что классификация не может быть тождественной филогении. Это утверждение вовсе не означает, что классификация непременно должна отражать что-то еще, кроме филогении (хотя некоторые авторы придерживаются именно такого мнения – см. I.5.2.2 и I.5.2.3). Можно строить классификацию, отражающую только филогенетические связи (как это постулируется кладистами – см. I.5.2.1), но в любом случае классификация так же не тождественна филогении, как любое отражение не тождественно отражаемому объекту. В классификации мы с помощью придуманных нами обозначений отражаем те стороны филогении, которые нам удалось выявить; сама же филогения существует независимо от нашего сознания и независимо от того, можем мы ее выявить или нет.
Различные авторы предлагают и отстаивают разные принципы объединения животных в таксоны. В настоящее время наиболее известными являются кладистический и традиционалистический принципы, именно они обсуждаются ниже. Кроме этого, можно назвать фенетический, или нумерический принцип систематики, который недолго использовался узким кругом авторов и себя не оправдал (см. выше, I.4.2.6.3.Б) и градистический принцип, малопопулярный среди современных систематиков (см. I.5.2.3). В то время как по поводу принципов выделения таксонов существуют разные мнения, термины, характеризующие таксоны по их отношению к филогенетическому древу, понимаются однозначно – это, прежде всего, термины голофилия, парафилия и полифилия.
I.5.1. Понятия моно-, поли-, пара- и голофилии
В зависимости от филогенетической природы таксонов их разделяют на монофилетические и полифилетические, либо на голофилетические, парафилетические и полифилетические.
I.5.1.1. История вопроса
Термины «монофилия» и «полифилия» были введены Геккелем в XIX веке и с тех пор широко употребляются, но при этом долгое время не имели четких определений; в результате этого разными авторами предложены различные определения этих понятий, иногда противоречивые. Было широко принято определение монофилетического таксона как «происходящего от одного общего предка» и полифилетического таксона, как «происходящего от нескольких разных предков». Однако если исходить из того, что все живые организмы произошли от общего предка, то определение монофилии как «происхождение от одного общего предка» надо считать явно недостаточным, так как в этом случае «монофилетическим» окажется любой таксон. Хенниг дал свое строгое определение понятия монофилия (см. ниже определение голофилии, или монофилии по Хеннигу), но при этом он сузил значение термина «монофилия» по сравнению с тем, как его интуитивно понимает большинство авторов. Поэтому были предложены другие, альтернативные определения.
Некоторые авторы [Simpson 1961, Mayr 1969 (русское издание – Майр 1971)] определяли монофилетический таксон как таксон, происходящий «по одной или более линиям от одного непосредственно предкового таксона того же самого или более низкого ранга». Очевидно, что такое определение абсолютно неприемлемо, поскольку определяющим в нем является искусственное понятие ранга таксона (см. ниже, I.5.4.2) и, таким образом, само понятие монофилии становится искусственным. При таком определении можно любой набор видов сделать «монофилетическим» таксоном, если приписать ему соответствующий ранг.
Например, если объединить насекомых с различными независимо возникшими сосущими ротовыми аппаратами в один таксон Haustellata Clairville 1789, то согласно общепринятому представлению о моно- и полифилии, таксон Haustellata является явно полифилетическим; однако если одновременно с этим всех прочих насекомых объединить в таксон Mandibulata Clairville 1798, и придать таксонам Haustellata и Mandibulata равные ранги (например, считать их обоих подклассами, как это делалось в некоторых работах конца XVIII – начала XIX века) (см. главу VI: Pterygota: «Классификации» I), то, согласно определению Симпсона, таксон Haustellata из полифилетического превращается в «монофилетический», хотя его состав и наши представления о его происхождении никак не изменились.
Приемлемые определения монофилии и полифилии, которые, с одной стороны, являются строго сформулированными, а с другой стороны, соответствуют традиционному пониманию этих терминов, были предложены Эшлоком (Ashlock 1971) (см. ниже). Одновременно с этим, чтобы не путать широкую монофилию и монофилию по Хеннигу, Эшлок предложил вместо монофилии по Хеннигу использовать новый термин голофилия. Термин парафилия был введен Хеннигом и всегда употребляется только в своем исходном значении (см. ниже).
I.5.1.2. Определения
Все таксоны делятся на монофилетические (в широком смысле слова) и полифилетические.
1. Монофилетический таксон (monophylum). Монофилетическим в широком смысле слова называется таксон, включающий в себя предка, общего для всех членов этого таксона, а также все филогенетические ветви, идущие от этого предка к каждому члену этого таксона. Под общим предком имеется в виду хотя бы самый ближайший из тех предковых видов, от которых произошли все прочие виды данного таксона. Поскольку общий предок таксона и другие предковые формы обычно в современной природе отсутствуют и исследователям неизвестны, будет точнее сказать, что предок не помещен в состав монофилетического таксона, а удовлетворяет диагнозу этого таксона.
Монофилетические (в широком смысле слова) таксоны делятся на голофилетические (или монофилетические в узком смысле слова) и парафилетические.
1.1. Голофилетический таксон (holophylum) – это то же, что строго монофилетический таксон, то есть монофилетический по Хеннигу. Голофилетическим является таксон, который не только включает общего предка и все филогенетические ветви, идущие от этого предка к каждому члену этого таксона, но также включает всех потомков этого предка. На рис. 1.2A показан голофилетический таксон b+c+d. Среди признаков, характеризующих голофилетический таксон, имеются аутапоморфии данного таксона, являющиеся при этом синапоморфиями всех видов, входящих в этот таксон. Действительно, апоморфия, возникшая у общего предка голофилетического таксона, может сохраниться у всех его потомков (т.е. во всем этом таксоне), а поскольку никакие потомки этого предка не находятся за пределами данного таксона, эта апоморфия имеется только в данном таксоне. Из определения явствует, что голофилетический таксон не может быть предковым для какого-либо другого таксона, поскольку любые формы, произошедшие от членов голофилетического таксона в результате эволюционных преобразований, включаются в этот же таксон. Если на филогенетической схеме обвести замкнутой линией голофилетический таксон, то эта линия пересечет филогенетическое древо только в одном месте – на входе в таксон. В слове голофилия «holo-» (от греческого ολος – целый) означает, что таксон образован целой ветвью филогенетического древа; эта ветвь отсечена от остального древа лишь в одном месте (являющимся основанием этой ветви) и включается в рассматриваемый таксон со всеми своими ответвлениями.
1.2. Парафилетический таксон (paraphylum). Парафилетическим называется таксон, включающий в себя предка, общего для всех членов этого таксона, и все филогенетические ветви, идущие от этого предка к каждому члену этого таксона, но включающий не всех потомков этого предка. Парафилия является разновидностью монофилии в широком смысле слова, но она отлична от монофилии по Хеннигу, т.е. от голофилии. В отличие от голофилетического, парафилетический таксон является предковым для другого таксона (или нескольких других таксонов) и отличается от него (или от них) лишь плезиоморфиями. На рис. 1.2B показан парафилетический таксон a+b+d+e, являющийся предковым для таксона c. Если на филогенетической схеме обвести замкнутой линией парафилетический таксон, то эта линия пересечет филогенетическое древо более чем в одном месте – один раз на входе в таксон и еще один или несколько раз на выходе (или выходах) из него.
2. Полифилетический таксон (polyphylum). Полифилетическим называется таксон, не включающий в себя предка, общего для всех членов данного таксона, либо не включающий в себя какие-либо участки филогенетических ветвей, соединяющих общего предка таксона с каким-либо из его членов (точнее сказать, эти предки не удовлетворяют диагнозу таксона). Признаки, на которых основан диагноз полифилетического таксона, возникали в эволюции независимо несколько раз. На рис. 1.2C показан полифилетический таксон a+b. Если на филогенетической схеме обвести замкнутой линией полифилетический таксон, то эта линия пересечет филогенетическое древо более чем в одном месте – не менее двух раз на разных входах в таксон.
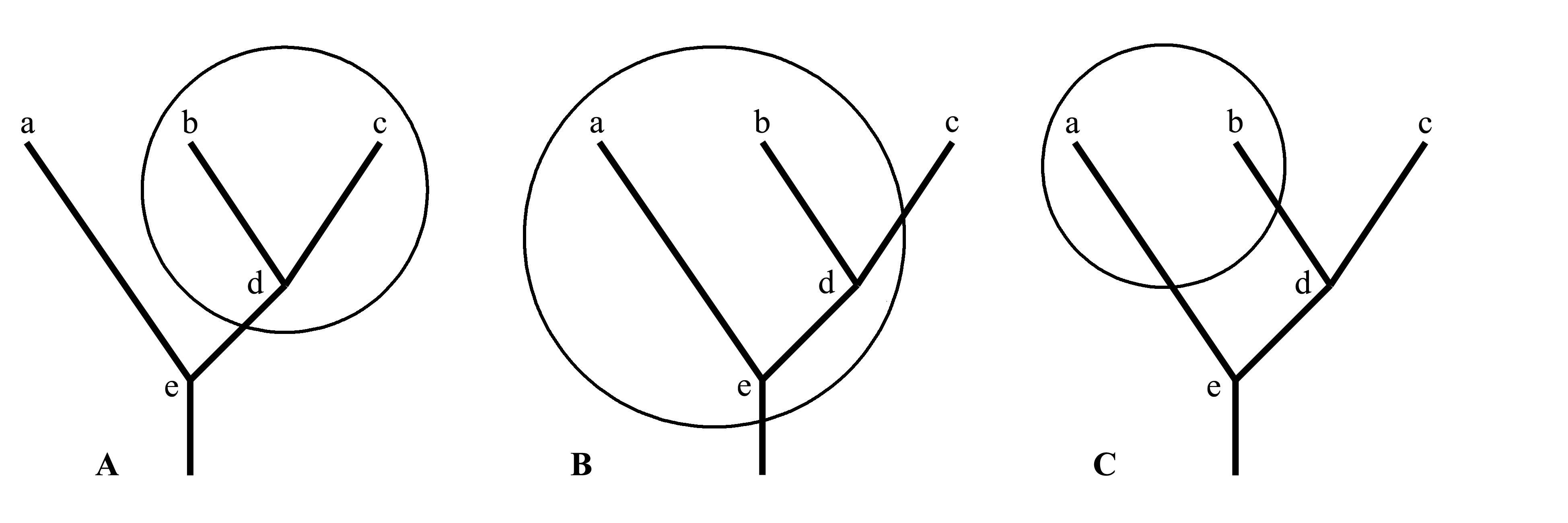
Рис. 1.2.
Три варианта выделения крупного таксона
в одной и той же группе животных, состоящей
из трех современных таксонов (a, b и c)
и предковых форм (d и e).
Филогенетические связи показаны жирной
линией, обсуждаемый таксон обведен окружностью:
A – голофилетический таксон (b+c+d); B –
парафилетический таксон (a+b+d+e); C – полифилетический
таксон (a+b).
Чтобы избежать оговорок «в широком смысле» и «в узком смысле» можно вообще не использовать двусмысленный термин монофилия, а обходиться терминами голофилия, парафилия и полифилия, каждый из которых является вполне однозначным.
I.5.1.3. Разъяснение понятий голо-, пара- и полифилия
Совсем кратко определения этих понятий можно выразить схемой:
|
Рассматриваемый таксон: |
включает |
включает |
|
включает общего предка |
голофилетический |
парафилетический |
|
не включает общего предка |
полифилетический |
|
Иногда неверное понимание этих терминов приводит к недоразумениям. Здесь мы обратим внимание на те моменты, которые иногда вызывают трудности в понимании и употреблении терминов голофилия, парафилия и полифилия.
Во-первых, следует помнить, что эти термины используются только применительно к таксонам, а не к организмам и не к филогенетическим ветвям. Поэтому прежде, чем обсуждать, является ли данный таксон голо-, пара- или полифилетическим, необходимо четко очертить этот таксон, т.е. договориться о том, какие виды в него включены, а какие нет. Обычно для установления границ таксона используется диагноз: в диагнозе перечисляется определенный набор признаков, и при этом предполагается, что если животное по своим признакам соответствует диагнозу данного таксона, то оно входит в этот таксон, а если не соответствует – то не входит. Если границы таксона не установлены, то понятия «включает» – «не включает» (предка или всех потомков) теряют смысл и определения становятся непригодными.
Установление границ таксона – процедура искусственная, как и сам таксон. После того, как таксон с его границами нами искусственным образом установлен, мы можем задать вопрос, насколько этот таксон является естественным, т.е. насколько он соответствует естественному объекту – филогенетической ветви, или иными словами, является ли он голофилетическим, парафилетическим или полифилетическим. Если изменить границы таксона, то изменится и ответ на вопрос, является ли он голо-, пара- или полифилетическим. Здесь мы считаем, что если мы изменили границы (циркумскрипцию) таксона, то мы перестали рассматривать прежний таксон, а рассматриваем уже другой таксон. Однако при использовании ранговой номенклатуры (см. ниже, раздел I.6), наоборот, таксон считается остающимся самим собой при изменении его границ; поэтому в соответствии с правилами номенклатуры одно и то же название присваивается таксонам с разными границами, что иной раз приводит к недоразумениям.
Например, род Phryganea Linnaeus 1758 исходно включал различных амфибиотических насекомых (ручейников и веснянок) и был полифилетическим таксоном; ныне таксон такой циркумскрипции никем не признается; однако в соответствие с правилами зоологической номенклатуры, родовое название Phryganea L. сохранено и сейчас применяется для таксона гораздо более узкой циркумскрипции, который является голофилетическим.
Во-вторых, ответ на вопрос, является ли данный таксон голо-, пара- или полифилетическим, будет однозначным только в соответствии с определенной гипотезой о филогении данной группы организмов.
Например, если мы примем гипотезу о родстве Onychophora, Myriapoda и Hexapoda (см. выше, схему в разделе I.4.1.5), то таксоны Gnathopoda, Euarthropoda и Mandibulata будут представляться нам полифилетическими, а таксон Ceratophora – голофилетическим; если же мы возьмем на вооружение гипотезу о родстве Eucrustacea, Myriapoda и Hexapoda (см. там же), то мы будем думать, что таксоны Gnathopoda, Euarthropoda и Mandibulata являются голофилетическими, а таксон Ceratophora – полифилетический. В действительности в отношении каждого таксона только один ответ является правильным, и задача биологов состоит в том, чтобы найти его, реконструируя филогению.
В-третьих, понятия голофилетический, парафилетический и полифилетический таксон являются абсолютными, в отличие от понятий апоморфный и плезиоморфный признак, которые являются относительными. Действительно, признак может быть апоморфным только по отношению к альтернативному ему признаку, который относительно его является плезиоморфным: признак B может быть апоморфным относительно признака A, но плезиоморфным относительно признака C (см. I.4.1.1). Таксон же является голофилетическим, парафилетическим или полифилетическим не относительно того или иного другого таксона, а вообще говоря. В определениях голо-, пара-, и полифилии фигурируют только естественные объекты «предок» и «потомки» и не упоминаются иные таксоны, кроме того таксона, к которому применяется определение. Например, если какие-либо потомки одного из членов таксона N не включены в таксон N, то таксон N называется парафилетическим независимо от того, как мы сгруппируем в таксоны этих потомков, не включенных в таксон N.
В-четвертых, понятия голо-, пара- или полифилетический таксон являются независимыми не только от классификации других таксонов, но и от признаков. Поскольку циркумскрипция (т.е. границы) таксона задается, как правило, с помощью признаков (образующих диагноз таксона), а филогения реконструируется на основе апоморфий (т.е. опять же признаков), создается неверное впечатление, что в основе всех этих понятий лежат признаки. Однако, как писал Линней, «не признак определяет род, но род определяет признак» – иными словами, признаки лишь помогают нам разобраться в этих понятиях, но не определяют их. Филогенетические связи существуют в природе независимо от того, сможем ли мы их реконструировать с помощью кладистического анализа (т.е. анализа признаков) или нет. Точно так же, если таксон включает своего общего предка и всех его потомков, этот таксон является голофилетическим, независимо от того, входят ли в его диагноз аутапоморфии или нет; если в его диагноз входят аутапоморфии, они являются для нас доказательством его голофилии; если аутапоморфии неизвестны или отсутствуют, мы не знаем, является ли данный таксон голофилетическим, но от этого нашего незнания он не перестает быть голофилетическим. Наличие в диагнозе таксона апоморфий – это не определение голофилии, а лишь способ ее выявления; но, поскольку этот способ выявления голофилии является единственным известным, некоторые ошибочно думают, что это и есть определение.
В-пятых, следует уточнить, что именно в определениях моно- и полифилии понимается под словом «предок». Каждый организм имеет множество предков, цепь которых восходит к нему от самого момента зарождения жизни на Земле. В свою очередь, каждый таксон имеет множество общих предков, в число которых входит вся цепь предков от зарождения жизни до первой из дивергенций, приведшей к разделению каких-либо из членов рассматриваемого таксона. Так что ни один таксон не может включать в себя всех своих предков. В определении монофилии (а также, соответственно, голофилии и парафилии) говорится, что таксон включает в себя какого-нибудь, хотя бы только самого ближайшего, из предков, общих для всех членов данного таксона. Соответственно, полифилетический таксон не включает ни одного из предков, общих для всех членов данного таксона.
I.5.2. Разные подходы к классификации
I.5.2.1. Кладизм, или филогенетическая систематика
Здесь и далее слово «кладизм» применяется к научно обоснованным принципам классификации, а не к формальным нумерическим и матричным методам, широко распространенным в современной биологической литературе (об их несоответствии научным принципам – см. выше, I.4.2.6).
Кладизм является единственным последовательным и четко сформулированным принципом построения классификации. Однако его положения выглядят парадоксальными, в связи с чем некоторые авторы его не признают. Как будет показано ниже (см. I.5.3), невозможно построить классификацию, которая бы полностью соответствовала требованиям кладизма, но все развитие систематики является приближением к такой классификации.
Принципы кладизма были сформулированы в книге Вилли Хеннига (Hennig 1950), причем автор называл эти принципы филогенетической систематикой. Словосочетание «филогенетическая систематика» не вполне удачно, так как подобные словосочетания (а именно «систематическая филогения» и «филогенетическая классификация») употреблялись в классических работах Геккеля в ином значении (Haeckel 1896, 1898). Термин «кладизм» образован от греческого слова κλαδος – ветвь. В кладистической систематике таксонами являются ветви филогенетического древа; можно также сказать, что кладистическая классификация – это классификация филогенетических ветвей, а не каких-либо других природных объектов.
Основную суть кладизма можно сформулировать очень кратко: все таксоны должны быть голофилетическими (т.е. монофилетическими по Хеннигу). Не допускается присутствие в классификации ни полифилетических, ни парафилетических таксонов.
Согласно этому принципу иерархия таксонов оказывается однозначно соответствующей иерархии ветвления филогенетического древа. Благодаря этому устраняется произвол в построении классификации. Действительно, если все таксоны должны быть только голофилетическими, не остается возможности выбора между разными способами разделения филогенетического древа на таксоны: во всех случаях граница между таксонами наиболее высокого ранга должна проходить по наиболее раннему по времени ветвлению рассматриваемой части филогенетического древа. Принцип кладизма влечет за собой важные следствия.
1) Если все таксоны являются голофилетическими, то никакие таксоны не могут быть материнскими (предковыми) и дочерними, а могут быть только сестринскими или не сестринскими.
Действительно, всякий таксон, предковый другому (не подчиненному ему) таксону, является парафилетическим по определению, и, следовательно, не должен существовать. Здесь не следует путать понятия предок и предковый таксон: существование предков признается кладистами так же, как оно признается и всеми прочими эволюционистически мыслящими биологами (лишь люди с крайне экстравагантными взглядами думают, что живые организмы могут возникать не от своих родителей, а как-то иначе). В отличие от предка, предковый таксон мы можем создать или не создавать; приверженцы кладистической систематики считают, что его создавать не следует.
2) Не все виды могут быть отнесены к таксонам всех рангов. Действительно, на рис.
1.3 вид с, являющийся общим предком и для
бабочек (Lepidoptera), и для ручейников (Trichoptera), не может быть отнесен ни к
бабочкам (так как в этом случае таксон
Lepidoptera стал бы парафилетическим), ни к
ручейникам (так как в этом случае парафилетическим стал бы таксон
Trichoptera). Для этого вида с также нельзя установить отдельный надвидовой таксон, противопоставляемый
Lepidoptera и Trichoptera, так как тогда это был бы парафилетический таксон. Так что общий предковый вид бабочек и ручейников не относится ни к
Lepidoptera, ни к Trichoptera, но вместе с бабочками и ручейниками относится к объединяющему их таксону
Amphiesmenoptera.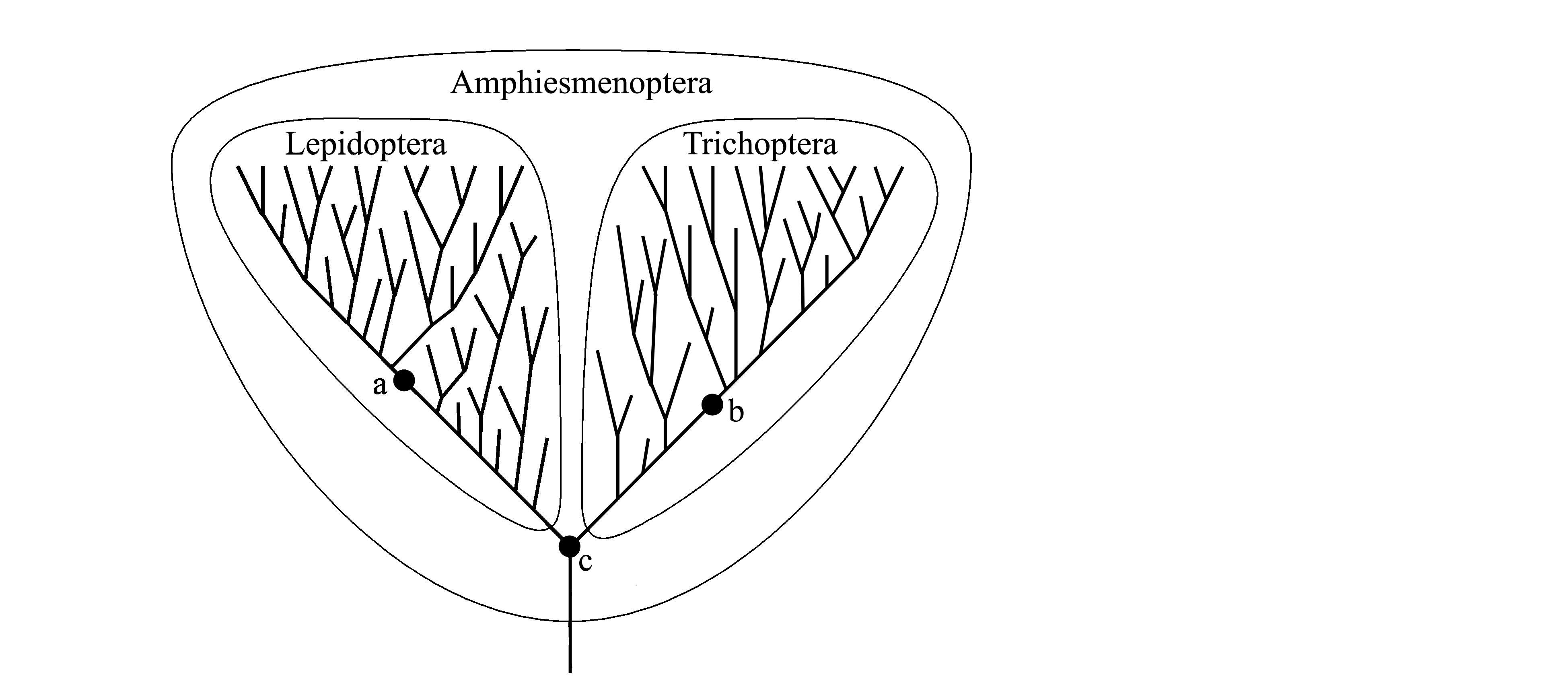
Рис. 1.3. Границы таксонов Lepidoptera, Trichoptera и
Amphiesmenoptera в кладистической систематике.
Жирной линией условно показаны
филогенетические ветви; тонкой линией
обведены границы таксонов; жирными точками
показаны следующие предковые виды: a –
общий предок всех современных Lepidoptera; b –
общий предок всех современных Trichoptera; c –
общий предок Lepidoptera и Trichoptera.
Если не следовать кладистическому принципу, а устанавливать парафилетические таксоны так, чтобы все виды включить в таксоны всех рангов, то оказывается неясным, где следует проводить границы между таксонами. Действительно, поскольку на филогенетическом древе естественных границ между таксонами нет (так как все древо представляет собой неразрывную ветвящуюся цепь поколений – рис. 1.1), в каждом конкретном случае трудно договориться, где должна проходить искусственная граница между таксонами. В кладизме предлагается универсальное решение этого вопроса: граница между двумя сестринскими таксонами должна проходить по их общему предковому виду, причем сам предковый вид не попадает ни в один из этих таксонов.
О реальном применении принципов кладизма см. ниже, I.5.3.
I.5.2.2. Традиционализм, или эволюционная систематика
Традиционализм возник как противопоставление кладизму и ставит своей целью защиту и обоснование тех положений, которые традиционно существуют в систематике, но отвергаются кладизмом. Идеи традиционализма сформулированы, в частности, в книге Майра (1971) и др. В имеющихся на сегодняшний день классификациях живых организмов присутствуют не только голофилетические, но и заведомо парафилетические таксоны. По мнению традиционалистов, парафилетические таксоны не только могут, но и должны существовать наряду с голофилетическими. Что касается полифилетических таксонов, то по мнению традиционалистов, так же как и по мнению кладистов, их быть не должно. При этом утверждается, что естественный таксон должен прежде всего характеризоваться общими признаками, унаследованными от предка; такими признаками являются не только апоморфии (характеризующие голофилетический таксон), но и плезиоморфии (характеризующие парафилетический таксон). В соответствии с этим, в традиционализме существует понятие предкового таксона – то есть родственные таксоны одного ранга могут быть не только сестринскими, но и материнским-дочерним.
Если кладисты считают, что для однозначности классификации она должна строиться на основе одного параметра – последовательности ветвления филогенетического древа (кладогенеза), то, по мнению традиционалистов, классификация должна строиться на основе двух параметров – 1) кладогенеза (cladogenesis), т.е. последовательности ветвления филогенетического древа и 2) анагенеза (anagenesis), т.е. величины эволюционных изменений в каждой ветви [здесь термин анагенез употребляется в широком смысле, как его определил Дж. Хаксли (Huxley 1957)]. Это различие обычно графически изображают как на рис. 1.4.
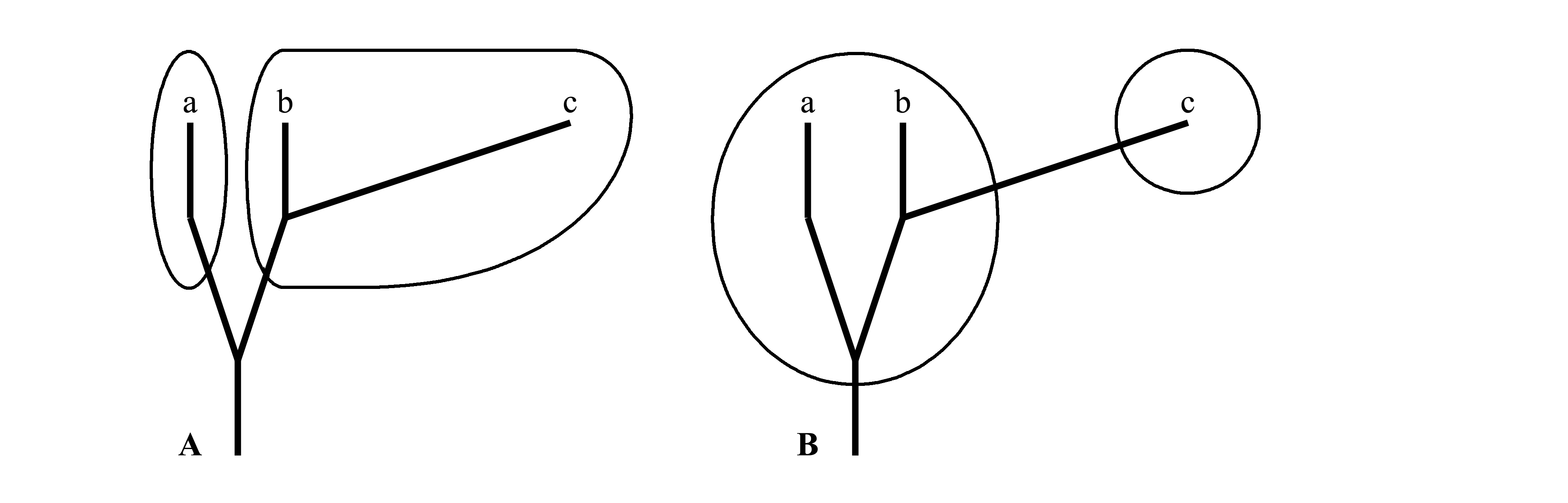
Рис. 1.4.
Два варианта разделения одной и той же
группы животных, состоящей из трех
современных таксонов (a, b
и c), на два крупных таксона.
Жирной линией показаны филогенетические
ветви; тонкой линией обведены границы
таксонов. Высота ветвей отражает
геологическое время; протяженность ветвей
по горизонтали призвана отражать величину
эволюционных изменений. А –
кладистическая классификация: каждый из
двух таксонов (таксон a и таксон b+c)
голофилетический. B –
традиционалистическая (эволюционистическая)
классификация: таксон a+b
парафилетический, таксон c
голофилетический.
На рас. 1.4А группа разбита на два таксона по кладистическому принципу, т.е. по наиболее ранней дивергенции, на рис. 1.4B та же группа разбита на два таксона иначе, в соответствии с традиционалистическим принципом, с учетом того, что ветвь c имеет большую анагенетическую составляющую. При этом неясно, как можно объективно определить величину эволюционных изменений (т.е. длину ветвей на рисунке). Традиционалисты не признают чисто количественной оценки степени различий (как это имеет место в фенетике); в отличие от фенетиков, они придают разным признакам разный вес и учитывают не только степень различия, но и последовательность ветвлений филогенетического древа.
Поскольку в традиционализме используется одновременно два параметра, неясно, какому из них в каких именно случаях следует отдать предпочтение. Поэтому в рамках традиционализма могут получаться разные классификации на основе одного и того же филогенетического древа: например, в случае на рис. 1.4 можно считать, что отличие c от a+b важнее последовательности ветвления, и на основании этого отдать предпочтение классификации «B», а можно, наоборот, считать это отличие менее важным, и на основании этого предпочесть классификацию «A».
О реальном применении принципов традиционализма см. ниже (I.5.3).
I.5.2.3. Градизм
В отличие от кладистической и традиционалистической школ систематики, каждая из которых претендует на то, чтобы быть единственной и служить основой для всей систематики, градизм обычно не рассматривается как всеобщий принцип построения классификации. Место градизма среди других подходов к систематике состоит в том, что некоторые авторы считают уместным существование градистических таксонов наряду с таксонами, установленными по другим принципам.
Можно образно сказать, что в отличие от кладистической классификации, где филогенетическое древо режется на таксоны вдоль, в градистической классификации оно режется на таксоны поперек. Такие таксоны, называемые также грады (gradus), представляют собой уровни организации и могут быть как голофилетическими, так и парафилетическими, и полифилетическими (рис. 1.5).
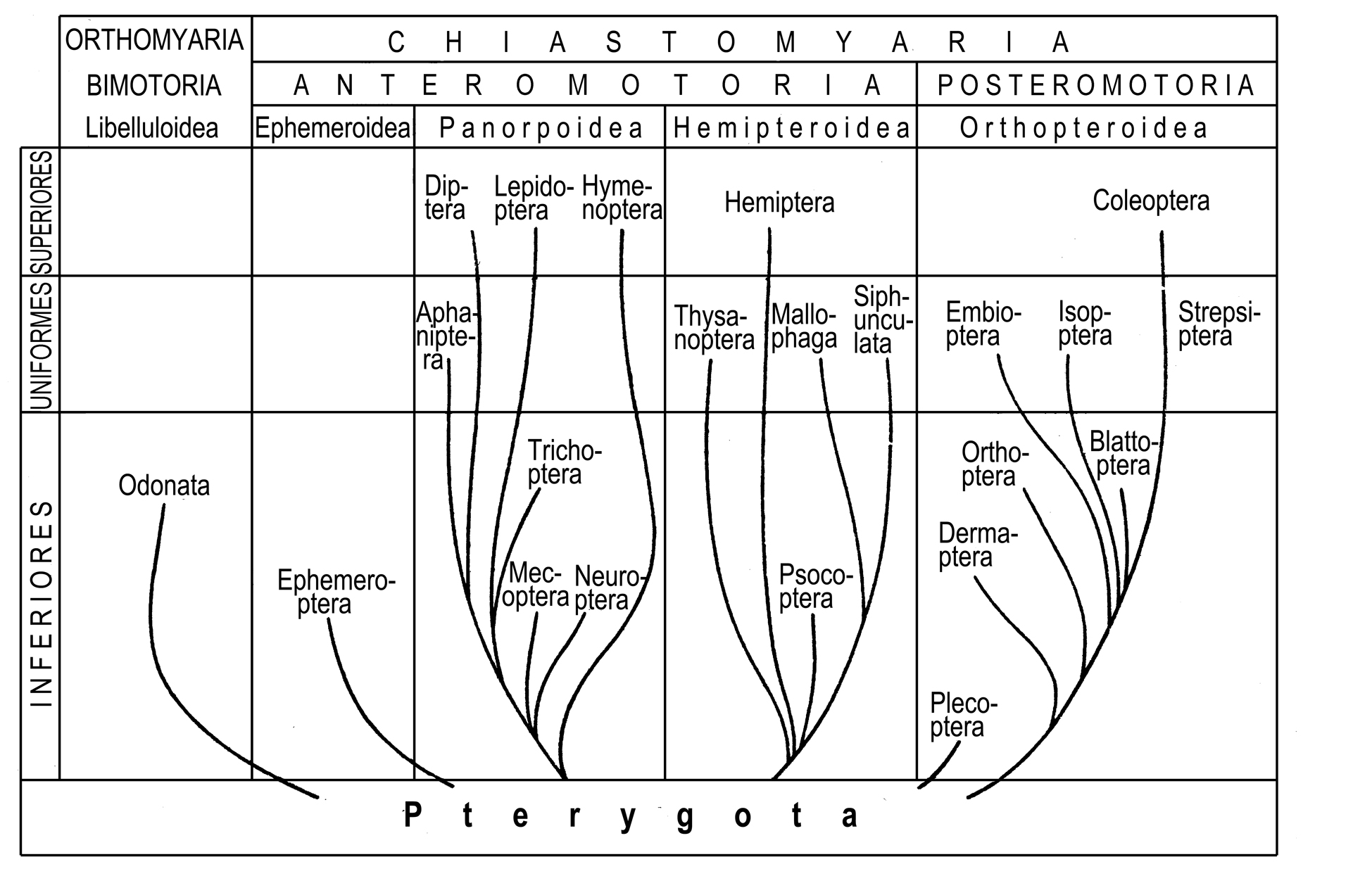
Рис. 1.5.
Градистическая классификация крылатых
насекомых по Б.Н. Шванвичу (Schwanwitsch 1946).
При этом нет ясного определения того, что следует считать разными уровнями организации. Можно произвольно выбрать любые из приобретенных в эволюции признаков, назвав их переходами на новые уровни организации; в зависимости от этого выбора получатся разные классификации на основе одного и того же филогенетического древа.
Наличие в градистических классификациях полифилетических таксонов определяется существованием конвергенции: диагнозы этих таксонов основаны на независимо возникших признаках. Если бы в конвергенциях существовала какая-нибудь закономерность, и наличие конвергенции по какому-либо признаку позволяло бы что-то прогнозировать, то классификация, основанная на конвергенциях, имела бы смысл. Однако в настоящее время такие закономерности неизвестны, и неясно, существуют ли они вообще. Поэтому градистический принцип построения классификации не имеет широкого признания, в отличие от кладистического (см. выше), где таксоны, будучи соответствующими филогенетическим ветвям, теоретически обоснованы и имеют явную прогностическую ценность.
I.5.3. Принципы классификации с точки зрения кладоэндезиса
Как было показано выше (см. I.5.2), разные авторы декларируют различные принципы построения классификации. Однако те принципы, которые декларируются, не всегда соответствуют принципам, которыми тот же автор руководствуется на самом деле при построении классификации.
Целесообразность существования в классификации голофилетических таксонов и нецелесообразность полифилетических признается как кладистами, так и традиционалистами. Разногласия вызывает вопрос о целесообразности существования парафилетических таксонов: кладисты их не признают, а традиционалисты признают.
Прежде чем перейти к обсуждению принципов кладоэндезиса, рассмотрим значение, которое имеют парафилетические таксоны.
I.5.3.1. Польза и вред парафилетического (предкового) таксона
Строго говоря, предком любого таксона у обоеполых организмов может быть только вид или более мелкая совокупность особей (подвид, раса, популяция), внутри которой осуществляется свободное перераспределение генетической информации; у организмов, не имеющих полового размножения, – это единичная особь. Однако вид, предковый для таксона, как правило, в современной природе отсутствует, в ископаемом состоянии неизвестен, и мы не имеем достаточно информации, чтобы теоретически реконструировать его с точностью до видовых признаков. Можно реконструировать интересующего нас предка с точностью до признаков таксона более высокого ранга (рода, семейства, отряда, класса, типа и т.п.).
Если в классификации допускается существование парафилетических надвидовых таксонов, то можно говорить не о предковом виде, а, соответственно, о предковом, или материнском (а, следовательно, парафилетическом) роде, семействе, отряде, классе, типе и т.п. При этом необходимо помнить, что в действительности предком таксона является не весь названный предковый таксон, а лишь один, неизвестный нам, но некогда существовавший вид этого таксона. Использование понятия предковый таксон иногда приводит к недоразумениям. Некоторым людям кажется, что, если какой-то надвидовой таксон называют предковым, то это значит, что разные виды этого таксона, каким-то необъяснимым образом «договорившись» друг с другом, эволюционировали в одном и том же направлении, образовав таким способом дочерний таксон надвидового ранга.
Например, если класс Hexapoda делится на подклассы Apterygota (первичнобескрылые) и Pterygota (крылатые) (см. главу VI: Hexapoda: «Классификации» I), то о подклассе Apterygota можно сказать, что он парафилетический и предковый для Pterygota или, что то же самое, Apterygota примитивнее, чем Pterygota. На основании этого утверждения некоторые люди всерьез думали, что у разных представителей Apterygota отрастали крылья, и они превращались в тех или иных представителей Pterygota. Кроме того, все виды, отнесенные к предковому, т.е. примитивному, таксону получают клеймо «примитивных»; дело доходит до того, что даже такую глубокую модификацию ротового аппарата, каковой является скрыточелюстность у Entognatha, некоторые авторы считали «примитивной» и предковой для открыточелюстного ротового аппарата других насекомых только на основании того факта, что насекомые, обладающие скрыточелюстным ротовым аппаратом, объединены с открыточелюстным предком крылатых насекомых в примитивный таксон Apterygota.
Утверждения «Apterygota дали начало Pterygota» или «Pterygota произошли от Apterygota» оказываются неправильно понятыми из-за того, что здесь одинаковые по форме слова Aptertygota и Pterygota (оба являющиеся названиями подклассов, оба во множественном числе) должны совершенно по-разному толковаться. Под словом «Apterygota» здесь следует понимать «какой-то из представителей таксона Apterygota», но ни в коем случае не «любой представитель Apterygota»; в то же время под словом «Pterygota» здесь же понимается «любой представитель Pterygota».
Это различие может быть понятным тому, кто устанавливает парафилетический таксон или высказывает подобные утверждения, но оно не всегда понятно тому, кто воспринимает эти утверждения.
Так, например, использование парафилетического понятия «обезьяна» в выражении «человек произошел от обезьяны» сделало это выражением любимой мишенью для нападок и издевок со стороны противников эволюционной теории и противников науки вообще. Некоторые люди, считающие себя эволюционистами, увидев в этом выражении одинаковые по форме слова «человек» и «обезьяна» считают, что если имеется в виду «всякий человек», то, значит, имеется в виду и «всякая обезьяна». В результате этого недоразумения научная теория о происхождении человека преобразуется в абсурдную версию о том, что если любой обезьяне дать в руки палку и создать ей еще какие-то загадочные условия, то в силу неких необъяснимых «законов эволюции» через много поколений ее потомки станут неотличимыми от людей. Естественно, что услышав такую версию, какой-нибудь не очень образованный, но критически мыслящий человек испытывает разочарование в науке. На вопрос «Утверждает ли современная наука, что человек произошел от обезьяны?» следует ответить, что современная наука этого утверждать не может, поскольку в ней нет термина, соответствующего слову «обезьяна». Действительно, в классификации К. Линнея (Linnaeus 1758) были и род Homo (человек), и род Simia (обезьяна); латинские названия обоих этих таксонов образованы по единой форме и, будучи названиями родовой группы, употребляются в единственном числе (см. I.6.3.1.4), так что переводятся именно как «человек» и «обезьяна». Пользуясь этой классификацией, мы могли бы вполне адекватно выразить современные эволюционные представления словами «человек (т.е. голофилетический род Homo) произошел от обезьяны (т.е. от парафилетического рода Simia)». Однако в современной классификации животных нет ни рода Simia в понимании Линнея, ни соответствующего ему по составу таксона какого-либо иного ранга. Для видов, относившихся Линнеем к родам Simia и Homo, Э. Геккель (Haeckel 1866) установил отряд Simiae, разделенный на подотряды Arctopitheci, Platyrrhinae и Catarrhinae (поскольку эти таксоны не являются родами, им присвоены нетипифицированные названия в форме существительных множественного числа; в отличие от родовых названий они пишутся не курсивом). В составе Catarrhinae принято выделять надсемейства Cercopithecoidea и Hominoidea; последнее делят на семейства Holobatidae и Hominidae; последнее делят на подсемейства Ponginae и Homininae; последнее делят на роды Gorilla, Pan и Homo. Здесь роду Simia в понимании Линнея соответствует совокупность всех видов таксона Simiae кроме рода Homo. В этой классификации каждый таксон является голофилетическим и не может быть предком какого-либо иного таксона, в том числе рода Homo: таксоны Simiae, Catarrhinae, Hominoidea, Hominidae и Homininae включают род Homo как составную часть, а таксоны Platyrrhinae, Cercopithecoidea, Holobatidae, Ponginae, Gorilla и Pan являются родственными, но не предковыми для человека.
Таким образом, вопрос о целесообразности парафилетических таксонов не имеет однозначного ответа: существование этих таксонов позволяет нам легче формулировать мысли о связях предок-потомок, однако легко сформулированная мысль не всегда легко и адекватно воспринимается.
I.5.3.2. Плезиоморфоны
В реально существующих классификациях, как правило, нет заведомо парафилетических таксонов (как нет и заведомо полифилетических), а есть только плезиоморфоны и заведомо голофилетические таксоны. Плезиоморфон (plesiomorphon) представляет собою таксон, характеризующийся только плезиоморфиями. Поскольку в характеристике плезиоморфона нет аутапоморфий, его голофилия не доказана и не предполагается. В то же время его нельзя обоснованно назвать парафилетическим, потому что парафилия также не доказана. Чтобы доказать парафилию таксона a+b на рис. 1.7A, надо найти синапоморфию между одной из его частей (b) и другим таксоном (c). Если такая синапоморфия будет найдена, таксон a+b можно с полным правом назвать парафилетическим; но одновременно с этим появится возможность расформировать его, объединив его бывшую часть b с таксоном c в новый голофилетический таксон b+c, характеризующийся новооткрытой апоморфией. Таким образом, плезиоморфон может быть фактически парафилетическим таксоном, но он существует в классификации только до тех пор, пока его парафилия не доказана. Некоторые из плезиоморфонов могут оказаться голофилетическими таксонами.
Термин «плезиоморфон» был введен недавно (Kluge 2004a) взамен неверно употреблявшегося термина «плезион». Термин «плезион» (plesion) исходно употреблялся не для того, чтобы указать статус таксона, а для того, чтобы обозначить место этого таксона в безранговой иерархической классификации; при этом плезион может быть как заведомо голофилетическим, так и парафилетическим (Patterson & Rosen 1977). В предыдущем издании (Клюге 2000) термин «плезион» ошибочно употреблялся для обозначения таксона, не являющегося заведомо голофилетическим – то есть для плезиоморфона. Некоторые авторы для обозначения плезиоморфона заключают название соответствующего таксона в кавычки (Patterson & Rosen 1977). Это неудачное обозначение, поскольку таким же способом обозначают неверное или сомнительное название таксона, независимо от статуса самого таксона.
I.5.3.3. Стремление классификации к кладистическому идеалу
По мере накопления знаний классификация все время меняется, при этом она стремится к такому идеалу, который представляет собой кладистическую классификацию; реально существующая классификация всегда приближается к этому идеалу, но никогда его не достигает.
Можно сформулировать следующее правило изменения классификации: классификация все время меняется так, что отношение суммарного объема (т.е. суммы видов) всех голофилетических таксонов к суммарному объему всех парафилетических таксонов увеличивается (под суммарным объемом здесь подразумевается сумма объемов всех, в том числе вложенных друг в друга, таксонов; т.е. суммарный объем всех надвидовых таксонов в несколько раз больше, чем общая сумма всех видов). Этот процесс хорошо прослеживается на примере изменения классификации в любой группе животных за любой период времени, как в доэволюционистический период, так и в эволюционистический период развития систематики.
Некоторые авторы полагают, что существует глубокое различие между кладистической и традиционалистической систематикой, поскольку кладизм допускает существование только голофилетических таксонов, а традиционализм – и голофилетических, и парафилетических. В действительности различия здесь могут быть только в декларируемых принципах, но не в результатах построения классификации.
Каждый кладистический таксон (который по определению должен быть голофилетическим, т.е. должен представлять собой филогенетическую ветвь) на самом деле может представлять собой не любую, а лишь такую филогенетическую ветвь, которая нам известна; нам может быть известна только та ветвь, которую удалось реконструировать с помощью филогенетического анализа (поскольку других методов выявления филогенетических ветвей неизвестно); филогенетический анализ обязательно предполагает наличие у реконструируемой ветви аутапоморфии, т.е. вполне определенного признака. Таким образом, в кладистической систематике каждый таксон обязательно характеризуется признаком, тогда как в природе далеко не каждая филогенетическая ветвь характеризуется признаком. Некоторые филогенетические ветви могут вообще не иметь апоморфий, и в этом случае мы на нынешнем этапе знаний не можем выявить такую ветвь, а, значит, не можем и придать такой ветви статус таксона. Если на рис. 1.6 на отрезке от E до D не произошло никаких эволюционных изменений, ветвь A+B не имеет никаких апоморфий, поэтому ее существование не может быть доказано, и, следовательно, для нее невозможно установить таксон.
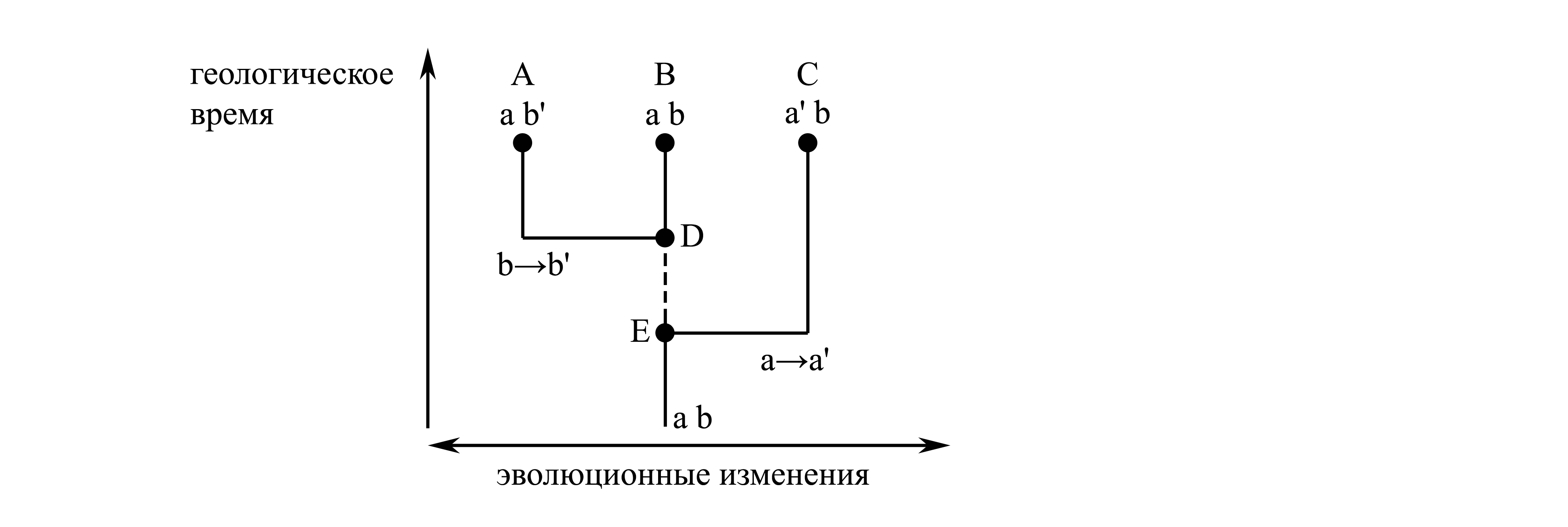
Рис. 1.6.
Филогенетические отношения, не поддающиеся
реконструированию. A, B и C –
современные таксоны; D и E –
предковые формы; a и a', b и b' –
признаки, среди которых a' и b' –
апоморфии. Прерывистой линией показан
участок филогенетического древа, не
поддающийся реконструированию.
Там, где не удается реконструировать филогению и образовать голофилетические таксоны, характеризующиеся аутапоморфиями, в рамках кладистической систематики создаются временные таксоны, характеризующиеся лишь плезиоморфиями – плезиоморфоны (см. I.5.3.2). Такой таксон, для которого аутапоморфии неизвестны, может оказаться парафилетическим или голофилетическим. Если в последствие доказывают парафилию этого таксона, его расформировывают, если доказывают его голофилию – сохраняют.
Так же на практике поступают и приверженцы традиционалистической систематики: признают существование парафилетического таксона, но только до тех пор, пока его парафилия не доказана. При этом традиционалисты декларируют, что должны существовать и голофилетические, и парафилетические таксоны – т.е. и синапоморфии, и симплезиоморфии являются естественными признаками и достойны того, чтобы на их основе объединять виды в надвидовые таксоны. Традиционалисты утверждают, что кладистическая систематика ущербна, так как отражает только кладогенез и не отражает анагенез (называемый также «филетической эволюцией», «степенью дивергенции» или «темпами эволюции»). Однако мы можем узнать о существовании кладогенеза (т.е. ветвления древа) только в том случае, если он сопровождается анагенезом (т.е. приобретением апоморфий), поскольку только кладистический анализ (т.е. анализ синапоморфий) позволяет выявить ветвления древа. Таким образом, утверждение, будто кладистическая систематика не отражает анагенез, неверно; в кладистической систематике анагенез отражен в той же мере, что и в традиционалистической.
Нередко можно услышать, что анагенез может иметь разную величину, и традиционалисты, в отличие от кладистов, при построении классификации принимают во внимание величину анагенеза. Однако никакого ясного определения, как оценивается величина анагенеза, не существует, и одно и то же эволюционное преобразование с равным основанием можно оценить и как очень большой анагенез, и как очень маленький.
Например, таксон крылатые насекомые (Pterygota) отличается от первичнобескрылых насекомых не только наличием сложно устроенных крыльев, но и преобразованием всего строения крылоносных сегментов груди (см. главу VII: Pterygota); поэтому можно было бы сказать, что возникновение этого таксона высокого ранга сопровождалось большим анагенезом (или ароморфозом, если пользоваться терминологией А.Н. Северцова). Но в таком случае становится непонятным, насколько большими следует считать те анагенезы, в результате которых среди Pterygota возникали многочисленные вторичнобескрылые виды и внутривидовые формы, поскольку у некоторых из этих форм не только утрачены крылья, но у грудь видоизменена так, что утратила все особенности, характерные для Pterygota.
В традиционалистической систематике для оценки величины анагенетической составляющей используется субъективное понятие «вес признака», но при этом принцип взвешивания признаков не объясняется. Фактически, как привило, наиболее весомым из изученных на данный момент признаков считают наиболее древнюю из апоморфий, свойственную наиболее крупной группе.
Например, если можно выбирать между классификациями I и II, выбирают ту из них, в которой таксону наиболее высокого ранга соответствует наиболее древняя из апоморфий.
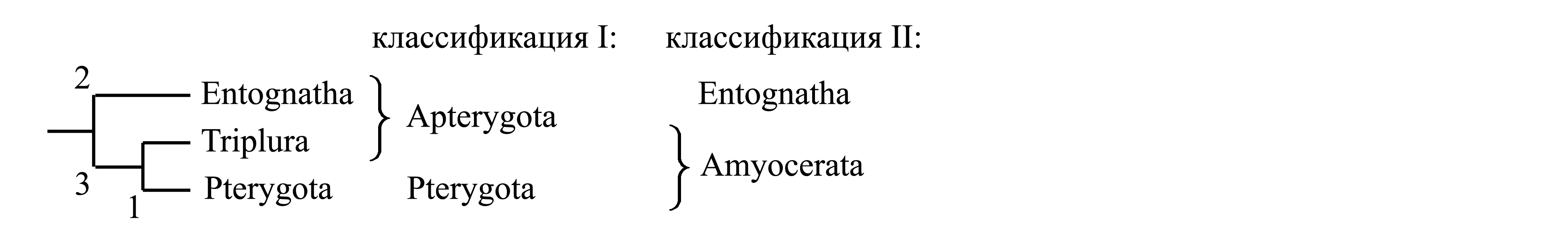
В данном случае в классификации I таксон наиболее высокого ранга – Pterygota, и ему соответствует набор аутапоморфий «1» – наличие крыльев и др. [см. главу VII: Pterygota (1)–(6)]. В классификации II таксон наиболее высокого ранга – Amyocerata, и ему соответствует аутапоморфия «3» – редукция мышц в жгутике антенны [см. главу VI: Amyocerata (1)]. Большинство систематиков, в том числе авторы, называющие себя традиционалистами, из этих двух классификаций выбирают классификацию II, в которой наиболее высокому по рангу таксону Amyocerata соответствует наиболее древняя из известных апоморфий «3», несмотря на то, что этот признак (упрощение строения антенн) является сам по себе малозаметным и не имеет никакого положительного значения для самих насекомых. Принимая классификацию II, тем самым отвергают классификацию I, в которой наиболее высокому по рангу таксону Pterygota соответствует менее древняя апоморфия «1», несмотря на то, что этот признак (возникновение крыльев) сыграл колоссальную положительную роль в филогенезе насекомых.
Здесь, как и во всех остальных случаях, вес признака определяется не свойствами самого признака, а местом признака на кладограмме. Таким образом, понятие «величина анагенеза» в рассуждениях систематиков представляет собой лишь производное понятия «кладогенез», а не особую объективную реальность.
Некоторые авторы пытаются оценить величину анагенетической составляющей путем подсчета признаков; эти попытки являются явно бессмысленными, так как морфологические признаки не дискретны и подсчитаны быть не могут (см. выше, I.4.2.6.3). В принципе можно объективно оценить величину анагенеза, подсчитав число уникальных замен нуклеотидов, но в настоящее время мы такими данными не располагаем.
Наши знания о филогении все время меняются по мере накопления новых фактов.
Например, в рассматриваемом здесь примере комплекс признаков «1» не был описан должным образом и не использовался в классификациях XVIII – начала XIX века (в частности, тогда было неясно, у каких насекомых крылья отсутствуют изначально, а у каких утрачены вторично, поэтому бескрылых Parasita Latreille 1796 рассматривали отдельно от крылатых и некоторых вторичнобескрылых насекомых, относившихся к Pterodicera Latreille 1802). Позже (в конце XIX века) признаки комплекса «1» стали лучше изучены, и это позволило реконструировать филогенетическую ветвь Pterygota; было также изучено строение челюстных карманов Entognatha (признак «2»), позволившее выявить этот таксон; но филогенетические отношения между Pterygota, Entognatha и Triplura (называвшимися тогда Ectotropha, Ectognatha, или Thysanura s.str.) оставались неясными. Основываясь на таких знаниях о филогении, таксон Hexapoda делили на два подчиненных таксона – плезиоморфон Apterygota и голофилетический таксон Pterygota.
Еще позже (Imms 1938) было выявлено распределение среди членистоногих признака «3» (наличие мышц лишь в первом членике антенн), что позволило более точно реконструировать филогенетическое древо и в дальнейшем на основании этого установить таксон Amyocerata Remington 1954 (или Ectognatha sensu Hennig 1953). При этом плезиоморфон Apterygota был расформирован, а в составе Hexapoda установлены два подчиненных голофилетических таксона – Entognatha и Amyocerata; ранее установленный голофилетический таксон Pterygota сохранился, став подчиненным таксоном в состав Amyocerata.
Другими иллюстрациями правила изменения классификации в направлении кладистического идеала по мере изучения признаков являются классификации, рассмотренные ниже в главах IV–X (см. Euarthropoda: «Классификации» I–II; Pterygota: «Классификации» I–VIII). В абстрактной форме это правило проиллюстрировано на рис. 1.7.
![]() Рис. 1.7.
Изменение классификации организмов в
процессе выявления новых апоморфий и
реконструирования филогении. Условные
обозначения: a–e – таксоны (a–c –
существующие, d–e – предковые и, как
правило, гипотетические); жирной линией
показано филогенетическое древо (сплошной
линией – реконструированные участки,
прерывистой линией – неизвестные участки);
тонкой линией обведены таксоны; 1, 2, 3 – номера
признаков, тонкая стрелка показывает
направление от плезиоморфии (белый
квадратик) к апоморфии (черный квадратик); holophyl. –
голофилетический таксон; paraphyl. –
парафилетический таксон; polyphyl. –
полифилетический таксон; толстые светлые
стрелки показывают изменение классификации,
построенной для одних и тех же организмов.
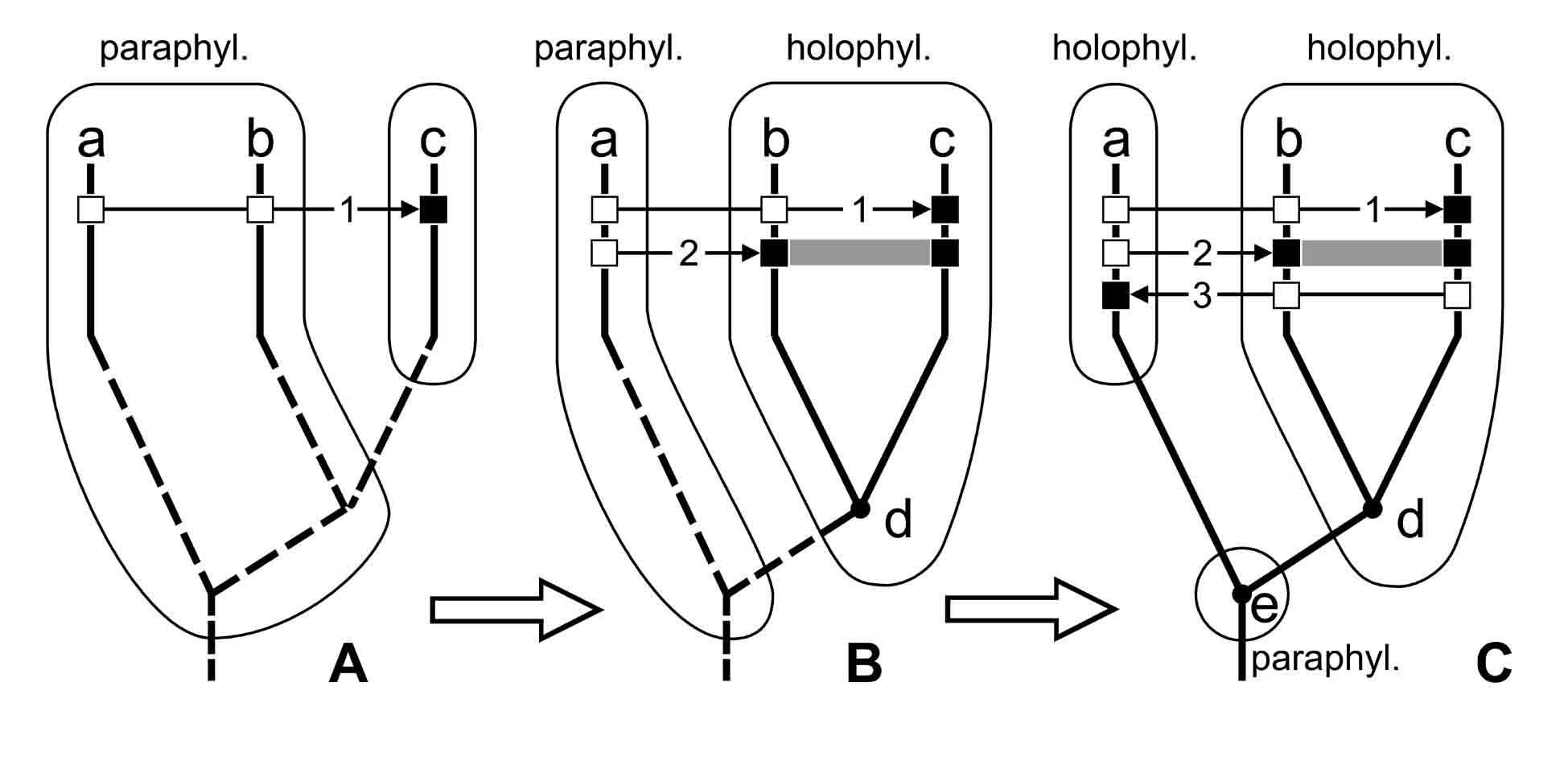
Рис. 1.7.
Изменение классификации организмов в
процессе выявления новых апоморфий и
реконструирования филогении. Условные
обозначения: a–e – таксоны (a–c –
существующие, d–e – предковые и, как
правило, гипотетические); жирной линией
показано филогенетическое древо (сплошной
линией – реконструированные участки,
прерывистой линией – неизвестные участки);
тонкой линией обведены таксоны; 1, 2, 3 – номера
признаков, тонкая стрелка показывает
направление от плезиоморфии (белый
квадратик) к апоморфии (черный квадратик); holophyl. –
голофилетический таксон; paraphyl. –
парафилетический таксон; polyphyl. –
полифилетический таксон; толстые светлые
стрелки показывают изменение классификации,
построенной для одних и тех же организмов.
Здесь парафилетический таксон a-b по мере накопления наших знаний о неизвестных ранее апоморфиях 2 и 3 сначала расформировывается с образованием меньшего парафилетического таксона (таксон a на рис. 1.7B), а затем (рис. 1.7C) заменяется голофилетическим таксоном, который характеризуется апоморфным признаком 3 и не включает гипотетический предковый вид e. По мере накопления знаний классификация стремится к такому состоянию, при котором парафилетический таксон сокращается до одного вида, являющегося общим предком для двух (или большего числа) других таксонов того же ранга (таковым может быть парафилетический таксон e на рис. 1.7C). Поскольку этот предок обычно неизвестен и является лишь гипотетическим, формально для него таксон не выделяется, и поэтому вся классификация состоит только из голофилетических таксонов. Такая классификация вполне удовлетворяет не только приверженцев традиционализма, но и приверженцев кладизма (так как в ней парафилетические таксоны формально отсутствуют).
Кладистический принцип можно рассматривать не как принцип, по которому должна быть построена классификация сейчас, а как идеал, к которому классификация должна все время стремиться в ходе ее изменения по мере накопления наших знаний об организмах.
Изменения классификации приближают ее к кладистическому идеалу, но этот идеал может быть достигнут только тогда, когда филогения будет полностью реконструирована. Филогения может быть полностью реконструирована только тогда, когда будет проведен кладистический анализ всех признаков. Для этого надо изучить все свойства организмов, т.е. полностью изучить все вопросы, поставленные перед биологией, и закрыть эту науку. Поскольку в обозримый период истории человечества этого не ожидается, классификация живых организмов все время будет меняться, приближаясь к кладистическому идеалу и не достигая его.
Это, к сожалению, делает классификацию нестабильной и создает много неудобств для исследователей. Нестабильность нынешней классификации неизбежна, поскольку классификация призвана быть естественной, а естественные свойства живых организмов, на которых она основана, во многом неизвестны и интенсивно изучаются. Если же отказаться от естественной классификации и создавать искусственную, то она не будет стабильной из-за своей субъективности.
I.5.3.4. Причины разногласий между кладистами и традиционалистами
Из всего сказанного в предыдущем разделе может создаться впечатление, что разногласий между кладистами и традиционалистами по поводу построения конкретных классификаций вообще быть не должно, а все споры могут сводиться только к теоретическому объяснению принципов систематики. Однако в действительности разногласия возникают именно по поводу того, какой должна быть классификация в той или иной группе организмов. Истинные причины этих разногласий состоят не в различиях между принципами кладизма и традиционализма, а в разном отношении к тому, насколько стабильной или нестабильной должна быть классификация.
Как было показано выше, естественная классификация животных стабильной в принципе быть не может и должна меняться по мере изменения представлений о филогении. Однако многие исследователи склонны проявлять оправданный консерватизм, стараясь не вносить изменений в классификацию до тех пор, пока необходимость этих изменений не станет очевидной. Приверженцы традиционалистической систематики склонны проявлять в этом отношении больший консерватизм, стараясь как можно дольше сохранять парафилетические таксоны. В отличие от них некоторые приверженцы кладистической систематики проявляют экстремизм, стараясь изменить классификацию сразу, как только появляется новая теория о филогении данной группы. Это вызвано тем, что согласно кладистическим принципам классификация уже сейчас должна быть построена как кладистическая (тогда как в действительности она может лишь приближаться к этому состоянию – см. выше). Подчас кладист, озабоченный задачей немедленно построить кладистическую классификацию, и в то же время не имея достаточно информации для реконструирования филогении, предлагает необоснованную филогенетическую гипотезу и на ее основе меняет классификацию.
Если эту необоснованную филогению можно опровергнуть, оппоненты опровергают ее, и, таким образом, спор концентрируется вокруг конкретных вопросов филогении. Однако кладист может предложить такую явно неверную филогенетическую гипотезу, которую в то же время нельзя быстро опровергнуть. Для этого существует два способа. Первый способ состоит в том, что филогению «реконструируют» с помощью ни на чем не основанного компьютерного метода (см. выше, I.4.2.6), так что сам автор этой «филогении» не может объяснить, в чем состоят его аргументы и ссылается при этом только на кем-то написанную компьютерную программу; поэтому и опровергнуть его аргументы невозможно. В этом случае некоторые традиционалисты, не имея возможности вести научный спор, и в то же время не имея возможности согласиться с предлагаемым изменением классификации, объясняют свою позицию как разногласие между кладизмом и традиционализмом.
Второй способ спровоцировать разногласия состоит в том, что кладист, желающий ускоренными темпами реконструировать филогению, впервые исследует какой-либо трудноизучаемый признак (в отличие от перетасовывания признаков на компьютере, такой род деятельности, безусловно, сам по себе является полезным и в любом случае вносит положительный вклад в науку). На изучение этого нового признака тратится много сил, времени и средств, в результате чего удается изучить этот признак для очень малого числа представителей огромных таксонов. Далее этот признак экстраполируется на всех неизученных членов этих таксонов, на основании чего строится кладограмма и меняется классификация.
Чтобы аргументировано опровергнуть эту филогению, нужно изучить этот же признак у других членов обсуждаемых таксонов, на что у оппонентов нет ни сил, ни времени, ни средств; в то же время сомнительность предложенной филогении не позволяет согласиться с новой классификацией.
Представим себе, что было бы, если бы люди не имели зрения и вынуждены были бы определять цвета предметов с помощью малодоступных дорогостоящих анализаторов. Тогда какой-нибудь исследователь, получивший редкую возможность поработать на таком анализаторе, выяснил бы, что комнатная муха имеет серый цвет, а жук-навозник и стрекоза-красотка имеют редкий в природе синий цвет; если синюю окраску расценить как синапоморфию, то, учитывая, что о цвете других насекомых ничего не известно, можно жесткокрылых и стрекоз объединить в таксон, противопоставляемый двукрылым.
Примерно такими же неправдоподобными, но в то же время неопровергаемыми выглядят некоторые современные гипотезы о филогении, основанные на малоизученных признаках. В таких случаях некоторые традиционалисты также пытаются объяснить свое неприятие новой классификации не разногласиями по поводу конкретных вопросов филогении, а разногласиями по поводу общих принципов систематики.
I.5.4. Относительные и абсолютные ранги таксонов
Выше рассмотрены те принципы, на основании которых виды животных группируются в таксоны. Далее рассмотрим принципы, на которых основана форма классификации. Эти принципы принято называть линнеевскими, хотя лишь некоторые из них были впервые предложены К. Линнеем, а некоторые взяты им из работ предшественников. Можно назвать следующие принципы линнеевской классификации, отличающие эту классификацию от других: (1) строгая иерархия классификации (иными словами, наличие относительных рангов таксонов); (2) наличие абсолютных рангов таксонов и (3) неравноценность рангов, т.е. наличие основных (обязательных) и неосновных (дополнительных) рангов и особый статус вида и рода. Из этих трех принципов первый обосновывается современными представлениями о филогенетической природе той «Естественной системы», которую призвана отражать линнеевская классификация; этот иерархический принцип с необходимыми поправками следует использовать и в современной систематике (см. ниже, I.5.4.1). Два других принципа до сих пор остаются широко признанными, однако они не отражают современных представлений о научной основе классификации животных, и их использование в систематике нецелесообразно (см. ниже, I.5.4.2 и I.5.4.3). Поэтому многие современные авторы стараются избегать абсолютных рангов. Однако до недавнего времени не было возможности полностью обойтись без рангов, так как с абсолютными рангами связана современная ранговая номенклатура таксонов. Теперь, с помощью новой безранговой иерархической номенклатуры надвидовых таксонов (см. ниже, I.6.4), появилась возможность записывать классификацию без абсолютных рангов, не вступая в противоречие с существующими правилами ранговой номенклатуры.
I.5.4.1. Строгая иерархичность классификации
Считается общепринятым, что классификация животных должна строиться по строго иерархическому принципу: каждый таксон входит в состав таксона более высокого ранга, причем один и тот же таксон не может входить в состав двух или более таксонов одинакового ранга. В этом отношении классификация видов живых организмов отличается от классификаций многих других природных объектов (химических элементов, веществ и др.). Например, в классификации химических веществ представляется вполне естественным, что аминокислоты относятся к кислотам и в то же время они относятся к органическим веществам, тогда как органические вещества в целом не относятся к кислотам, а кислоты в целом не относятся к органическим веществам. В противоположность этому, в классификации живых организмов совершенно недопустимо, чтобы например, какой-нибудь отряд относился к двум разным классам одновременно.
С давних времен система живых организмов создавалась не просто как удобная классификация, а как отражение объективно существующей в природе закономерности (как писал К. Линней, «не признак определяет род, но род определяет признак»). Однако лишь с развитием эволюционной теории стал в общих чертах понятен природный механизм, благодаря которому эта закономерность существует. Единство каждого таксона обеспечивается происхождением всех членов этого таксона от общего предка, а иерархия классификации обеспечивается тем, что увеличение числа видов происходит путем дивергенции (разделения одного предкового вида на два или несколько дочерних), тогда как образование одного вида от двух или нескольких предковых в большинстве случаев невозможно.
Здесь уместно обратить внимание на одну деталь, часто упускаемую из вида. Естественную иерархическую систему с более или менее четко очерченными видами и таксонами более высоких рангов образует не все филогенетическое древо, а лишь его фрагменты, доступные для нашего изучения: это современный срез филогенетического древа (т.е. совокупность всех современных организмов) и сохранившиеся ископаемые. Для этих доступных изучению объектов мы можем построить непротиворечивую иерархическую классификацию, где все надвидовые таксоны разделены естественными разрывами. Если же взять всю совокупность когда-либо существовавших организмов (что можно сделать лишь мысленно), то они образуют единое филогенетическое древо, т.е. непрерывную ветвящуюся цепь поколений, и не распадаются ни на изолированные виды, ни, тем более, на таксоны более высокого ранга (рис. 1.1). Если мы представим себе классификацию всех живших на Земле организмов, то в этой классификации границы таксонов окажутся совершенно искусственными: граница между соседними таксонами любого, даже самого высокого ранга, в каком-то месте обязательно пересечет цепь поколений таким образом, что мать окажется в одном таксоне, а ее родной ребенок в другом. К. Линней в «Философии ботаники» писал: «Искусственные классы замещают естественные, пока все естественные не открыты: тогда же, когда с открытием еще многих родов они будут выявлены, весьма трудно будет установить четкие границы классов».
I.5.4.2. Абсолютные ранги
Помимо того, что в линнеевской системе существуют относительные ранги (см. выше), в ней имеются также абсолютные ранги (такие как класс, отряд, род, вид и др.). В соответствии с этим в одной и той же классификации разными таксонами могут считаться не только группы, объединяющие разные наборы видов, но и идентичные по составу группы, различающиеся только абсолютным рангом.
Например, если отряд Protephemeroidea включает единственное семейство Triplosobidae, куда входит единственный род Triplosoba с единственным видом Triplosoba pulchella, то таксоны Protephemeroidea, Triplosobidae, Triplosoba и T. pulchella идентичны с биологической точки зрения, но они имеют разный ранг и поэтому формально считаются разными таксонами.
Если таксон объединяет несколько таксонов следующего ранга, он называется политипическим. Если таксон включает лишь один непосредственно подчиненный таксон, он называется монотипическим; монотипический таксон ничем, кроме абсолютного ранга, не отличается от непосредственно подчиненного ему таксона, то есть не имеет собственного биологического смысла. Понятие монотипии является чисто формальным: например, если несколько видов объединены в одно семейство и в один род, то это семейство будет считаться монотипическим; но если это же семейство с этим же набором видов разделить на несколько родов, то семейство будет считаться политипическим.
Общепринятые ранги, используемые в зоологии, перечислены в табл. I.3 и I.4.
Таблица
I.3. Ранги зоологических таксонов в убывающей
последовательности
|
Единственное |
Множественное |
Сокращение |
Русский |
Статус |
Источник |
|
imperium |
imperia |
imp. |
империя
|
доп. |
Linnaeus
1758 |
|
regnum |
regna |
regn. |
царство |
основной |
Linnaeus
1758 |
|
phylum |
phyla |
phyl. |
тип |
основной |
Haeckel 1866
(phylum) |
|
classis |
classes |
cl. |
класс |
основной |
Linnaeus
1758 |
|
legio |
legiones |
leg. |
легион |
доп. |
Latreille
1806 |
|
cohors |
cohortes |
coh. |
когорта |
доп. |
Latreille
1806 |
|
ordo |
ordines |
ord. |
отряд |
основной |
Linnaeus
1758 |
|
familia |
familiae |
fam. |
семейство |
основной |
Latreille
1802 (до этого использовался вместо
отряда) |
|
tribus |
tribus |
tr. |
триба |
доп. |
Cuvier 1817 (до
этого использовался для рангов выше
семейства или ниже рода) |
|
genus |
genera |
gen. |
род |
основной |
Linnaeus
1758 |
|
species |
species |
sp. |
вид |
основной |
Linnaeus
1758 |
|
varietas |
varietates |
var. |
разновидность |
доп. |
|
|
forma |
formae |
f. |
форма |
доп. |
|
Таблица
I.4. Общепринятые префиксы для образования
дополнительных рангов (в порядке от более
высоких к более низким)
|
Латинский |
Русский |
Примечание |
|
super- |
над- |
не
может использоваться для рода |
|
sub-
|
под- |
может
использоваться для всех рангов |
|
infra- |
инфра- |
в
зоологии не используется для рода и вида |
|
subter- |
субтер- |
в
зоологии не используется для рода и вида |
Из перечисленных в табл. I.3 рангов regnum (царство), classis (класс), ordo (отряд), genus (род) и species (вид) использовались в качестве обязательных в работах К. Линнея (Linnaeus 1736–1796). Позже в качестве обязательных добавились ранги familia (семейство) и phylum (иногда называвшийся также typus – тип).
В 10-м издании книги К. Линнея «Система природы» (Linnaeus 1758), ныне официально принятом в качестве отправной точки зоологической номенклатуры (см. ниже, раздел I.6), классификация выглядит следующим образом. Империя Natura (природа) делится на 3 царства – Animalia (животные), Vegetabilia (растения) и Lapides (минералы). Каждое царство делится на классы; в частности, царство Animalia делится на 6 классов – Mammalia (млекопитающие), Aves (птицы), Amphibia (сборная группа позвоночных), Pisces (рыбы), Insecta (членистоногие) и Vermes (прочие животные). Каждый класс делится на отряды; в частности, класс Insecta делится на 7 отрядов – Coleoptera, Hemiptera, Lepidoptera, Neuroptera, Hymenoptera, Diptera и Aptera. Каждый отряд делится на роды. Отряд Coleoptera делится на 25 родов; среди них, помимо 22 родов, ныне относимых к жесткокрылым, имеются роды Forficula (уховертки), Blatta (тараканы) и Gryllus (прямокрылые в широком смысле). Отряд Hemiptera (соответствующий современному таксону Condylognatha) делится на 7 родов. Отряд Lepidoptera (чешуекрылые) делится на 3 рода. Отряд Neuroptera (не соответствующий ничему в современных классификациях) делится на 6 родов – Libellula (стрекозы), Ephemera (поденки), Phryganea (ручейники и веснянки), Hemerobius (двухоботные, вислокрылки, сеноеды и крылатые особи термитов), Panorpa (скорпионницы) и Raphidia (верблюдки). Отряд Hymenoptera (перепончатокрылые) делится на 8 родов. Отряд Diptera (двукрылые) делится на 10 родов. Отряд Aptera (не соответствующий ничему в современных классификациях) делится на 14 родов – Lepisma (трехвостки), Podura (ногохвостки), Termes (бескрылые особи термитов), Pediculus (вши и пухоеды), Pulex (блохи), Acarus (клещи), Phalangium (сенокосцы, телифоны и фрины), Aranea (пауки), Scorpio (скорпионы), Cancer (раки и крабы), Monoculus (некоторые ракообразные и др.), Oniscus (мокрицы и др.), Scolopendra (губоногие и др.) и Julus (двупарноногие). Каждый род делится на виды. В некоторых отрядах роды сгруппированы в таксоны промежуточного ранга. В частности, в отряде Aptera первые пять родов объединены в группу без ранга и без названия – шестиногие с обособленной головой; следующие шесть родов объединены в группу многоногих с необособленной головой, а последние три рода – в группу многоногих с обособленной головой. Некоторые роды подразделены на промежуточные таксоны, каждый из которых уже делится на виды. В частности, род Gryllus подразделен на таксоны без ранга Mantis (богомолы и палочники), Acrida, Bulla, Acheta, Tettigonia и Locusta (пять последних соответствуют прыгающим прямокрылым).
В дальнейшем менялась не только иерархия таксонов, постепенно приближаясь к более естественной (о принципе этого изменения см. выше), но менялись и абсолютные ранги таксонов. У некоторых из сохранившихся линнеевских таксонов ранги также сохранились без изменения до наших дней: это, например, классы Mammalia и Aves, отряды Lepidoptera, Hymenoptera, Diptera и др. У других сохранившихся до наших дней таксонов ранги сильно изменились: например, у Forficula, Libellula, Ephemera, Panorpa, Raphidia, Lepisma, Podura, Pediculus и Pulex ранг вырос от рода до отряда или выше; у Julus ранг вырос от рода до класса; у некоторых таксонов в составе Vermes ранг вырос от рода до типа.
Ранги являются привычным компонентом систематики; более того, в соответствии с Международным кодексом зоологической номенклатуры (см. ниже, раздел I.6), тем таксонам, которые подпадают под правила этого кодекса, узаконенные названия могут быть присвоены только в том случае, если этим же таксонам присвоены конкретные ранги.
Однако целесообразность использования рангов вызывает сомнения. Только один ранг – вид – имеет научно обоснованное определение: критерием биологического вида является наличие репродуктивной изоляции от других видов и отсутствие репродуктивной изоляции внутри вида (см. I.3). Значение всех остальных рангов сводится лишь к тому, чтобы показать иерархическую соподчиненность таксонов: таксон более высокого ранга делится на таксоны более низкого ранга. Некоторые авторы считают, что ранги должны нести еще какую-то научную информацию, при этом расходятся во мнении, какую именно. В соответствии с тем, какое значение хотят придать рангам, предлагают противоречащие друг другу критерии рангов: одни авторы предлагают связать ранги со степенью дивергенции таксонов, другие – с их возрастом.
Связать ранги со степенью дивергенции невозможно, потому что нет универсального критерия для оценки степени дивергенции.
Автор филогенетической систематики В. Хенниг считал, что если в филогенетической систематике относительные ранги таксонов (т.е. их положение в иерархии относительно друг друга) зависят от относительного времени дивергенции (т.е. от последовательности ветвления филогенетического древа – см. I.5.2.1), то абсолютные ранги таксонов (т.е. типы, классы, отряды и т.п.) должны зависеть от абсолютного (т.е. геологического) времени дивергенции. Для того, чтобы ранги таксонов насекомых, установленные по геологическому возрасту, в наибольшей степени соответствовали традиционно принятым рангам этих таксонов, Хенниг предложил следующее соответствие между рангами и временем обособления таксонов: класс – от кембрия до начала девона; отряд – от конца девона до конца перми; семейство – от начала триаса до конца раннего мела; триба – от начала позднего мела до конца олигоцена. Однако если использовать эту же шкалу для таксонов позвоночных, придется присвоить им ранги более низкие, чем ныне принятые, в частности таксон Mammalia (млекопитающие) придется считать не классом, а отрядом; некоторые другие таксоны придется повысить в рангах: в частности, некоторые таксоны, ныне считающиеся родами в составе класса Ostrapoda и известные начиная с ордовика, придется считать не родами, а классами. Применение этого критерия рангов наталкивается на большие трудности. Абсолютное время дивергенции можно установить только на основе палеонтологических данных. При этом можно достоверно установить, что дивергенция произошла не позже такого-то времени (если в отложениях этого времени найдены остатки организмов, возникших в результате данной дивергенции), но нет способа, который позволил бы достоверно установить, что данная дивергенция произошла не раньше какого-то определенного времени. Поэтому гипотезы о времени дивергенции часто имеют очень большой разброс. Для многих групп организмов вообще нет никаких ископаемых остатков, и для них этот критерий рангов не применим.
Поскольку нет общепринятых критериев для надвидовых рангов, в ныне существующих классификациях ранги, как правило, установлены произвольным образом. Ранги являются отчасти естественным, отчасти искусственным компонентом систематики: их относительные значения могут быть естественными, тогда как абсолютные значения являются искусственными. Если классификации различаются только абсолютными значениями рангов, между этими классификациями нет принципиальной разницы.
Использование рангов часто порождает неверное представление, будто любые таксоны одного ранга имеют нечто общее между собой. В результате этого в некоторых научных исследованиях таксоны одного ранга неверно используют в качестве сопоставимых элементов. Например, сравнивают фауны по числу общих родов или рассматривают филогению строго на уровне семейств. В действительности таксоны одного ранга являются сопоставимыми элементами, только если это сестринские таксоны в составе одного таксона более высокого ранга: например, роды одного семейства сопоставимы между собой, тогда как роды из разных семейств лишь формально считаются таксонами одного ранга, и их сопоставление неправомерно.
В главах III–X данной книги все таксоны рассматриваются без придания им абсолютных рангов, что оказалось возможным благодаря последовательному использованию безранговых номенклатур (см. ниже, I.6); перед характеристикой каждого таксона в рубрике «Ранг» указаны те ранги, которые чаще других приписывают этому таксону.
I.5.4.3. Неравноценность абсолютных рангов
Среди абсолютных рангов различаются основные и дополнительные (см. табл. I.3). Среди основных рангов (вид, род, семейство, отряд, класс, тип и царство) ранги вид и род имеют особый статус. Основные ранги являются обязательными, а дополнительные – необязательными.
Если в классификации иерархических уровней меньше, чем существует основных рангов, то используются все основные ранги, даже если в этом нет необходимости.
Например, если один вид Triplosoba pulchella не может быть помещен ни в какой ранее установленный отряд, для него устанавливаются новый монотипический отряд Protephemeroidea; в этом отряде формально устанавливается новое монотипическое семейство Triplosobidae и новый монотипический род Triplosoba (циркумскрипции и диагнозы которых совпадают с циркумскрипцией и диагнозом вида и отряда).
Таким образом, в классификации появляются избыточные таксоны и названия.
Если в классификации иерархических уровней больше, чем существует основных рангов, то используются все основные и необходимое количество дополнительных рангов. При этом присвоение тому или иному таксону основного или дополнительного ранга является произвольным.
Для примера возьмем классификацию насекомых, записанную без рангов, и припишем этим группам ранги двумя разными способами (в современной литературе используется гораздо больше разных способов расстановки рангов для этих таксонов):

Поскольку дополнительные ранги являются необязательными, многие авторы, воспроизводя классификацию, их опускают, а вместе с ними опускают и таксоны, которым эти ранги приписаны, и диагнозы этих таксонов. В результате этого в некоторых публикациях в составе типа членистоногих рассматривается класс Hexapoda, для которого дается развернутая характеристика, но при этом ничего не говорится о таксоне Amyocerata; в других работах, наоборот, дается развернутая характеристика класса Amyocerata, а таксон Hexapoda игнорируется. Это порождает ложное впечатление, будто речь идет о принципиально разных классификациях. Дело доходит до того, что некоторые люди всерьез думают, что кто-то отменил один из этих таксонов, признав его искусственным; возникновению такого рода недоразумений способствует использование рангового принципа номенклатуры (см. ниже, I.6.6.2).
I.5.4.4. Проблема рода
Особый статус ранга рода выражается в том, что, согласно линнеевскому принципу, закрепленному в Международном кодексе зоологической номенклатуры (см. ниже, I.6.3), вид может получить бинарное название только в том случае, если он хотя бы формально отнесен к какому-нибудь роду. Поэтому чтобы назвать вид, необходимо не только поместить его в какой-нибудь надвидовой таксон, но и выбрать, который из надвидовых таксонов, включающих этот вид, считать родом. При перемещении ранга рода с одного надвидового таксона на другой меняются названия видов (тогда как при изменении рангов других надвидовых таксонов могут меняться только названия этих таксонов, но не названия входящих в них видов). В то же время родовой ранг является наименее стабильным из всех, он чаще других перемещается с одного систематического таксона на другой.
Это противоречие вызвано тем, что в ходе развития систематики под одним и тем же словом «род» («genus») понимали три принципиально разные вещи. Следует различать аристотелевский род, линнеевский род и латрейлевский род.
Аристотелевский род является относительным понятием: это любая категория, объединяющая виды, в то время как Аристотелевский вид – это любая категория в составе рода. По Аристотелю виды объединяются в роды; если эти роды в свою очередь объединяются в род более высокого порядка, то они является видами по отношению к этому вышестоящему роду; соответственно, вид может быть подразделен на виды более низкого порядка, и тогда он является родом по отношению к ним. Таким образом, вся иерархия описывается двумя категориями – вид и род. Аристотель считал, что иерархия является фундаментальным свойством всей природы, так что аристотелевские вид и род были фундаментальными категориями.
Линнеевский род, в отличие от аристотелевского, является не относительны, а абсолютным. Линней называл родами не любые таксоны, объединяющие виды, а таксоны только одного иерархического уровня: для каждого вида лишь один вышестоящий таксон называется родом, а таксон, объединяющий несколько родов, называется не родом более высокого порядка, а иначе (например, отрядом). Таким образом, иерархия описывается не двумя относительными категориями (вид и род), а несколькими абсолютными категориями – вид, род, отряд, класс и др. В этой системе род оказался не фундаментальной категорией, а лишь одной из нескольких равноценных. Несмотря на то, что Линней изменил смысл слов «вид» и «род», он продолжал считать вид и род фундаментальными категориями и утверждал, что виды и роды более естественны, чем таксоны других рангов. Особый статус вида Линней видел в том, что внутривидовые таксоны эволюционируют, а виды и все таксоны выше вида неизменны; в настоящее время мы считаем это представление ошибочным, но если видом называется биологический вид (см. I.3)], то он действительно имеет особый статус, поскольку внутривидовые таксоны не имеют репродуктивной изоляции, а виды и все таксоны выше вида репродуктивно изолированы друг от друга. Что касается особого статуса рода, то Линней его никак не объяснил; видимо, представление о фундаментальности понятия «род» было просто механически перенесено из философии Аристотеля в систематику.
Бинарная номенклатура была разработана Линнеем с учетом того, что исходно в его системе роды представляли собой наиболее стабильные таксоны. Если в качестве рода была выделена естественная группа, этот род с его диагнозом и циркумскрипцией (т.е. границами) сохранялся при последующих изменениях классификации. Если открывали новые виды, соответствующие диагнозу ранее установленного рода, их помещали в этот род. Таким образом число видов в роде увеличивалось, но род оставался самим собой. При увеличении числа видов возникает необходимость их упорядочить, для чего приходится увеличивать число рангов и таксонов. Линней вводил дополнительные ранги, помещая их по мере необходимости как ниже рода, так и выше рода. Благодаря этому увеличение числа известных видов не мешало стабильности родов и не приводило к изменению бинарных названий ранее описанных видов. Лишь в случае, если в качестве рода была выделена искусственная группа, такой род расформировывался.
В линнеевской системе только таксоны основных рангов (класс, отряд, род и вид) имели единообразные названия, тогда как для других таксонов единого принципа номенклатуры не было: например, род Gryllus делился на безранговые таксоны с названиями в единственном числе – Mantis, Acrida, Bulla, Acheta, Tettigonia и Locusta, а род Cicada делился на безранговые таксоны с названиями во множественном числе – Noctilucae, Foliaceae, Cruciatae, Manniferae, Spumantes и Diflexae.
Латрейлевский род является абсолютным лишь в определенной классификации, но при изменении классификации он оказывается относительным. В конкретной классификации для каждого вида лишь один конкретный вышестоящий таксон называется родом (как в случае с линнеевским родом), но в разных классификациях ранг рода может приписываться разным таксонам, вышестоящим по отношению к этому виду. По мере увеличения числа известных видов ранг рода приписывается таксону все более и более низкого уровня. Циркумскрипция линнеевского рода может быть изменена только в том случае, если при его установлении если была допущена ошибка; изменение циркумскрипции латрейлевского рода является нормальным действием, не связанным с ошибками. Такая ситуация сложилась по следующим причинам.
Линней объединял роды животных и растений в отряды, которым дал преимущественно нетипифицированные названия. Ботаники того времени называли таксон между родом и классом либо отрядом – ordo (Jussieu 1789), либо семейством – familia (Adanson 1763), и давали некоторым из этих таксонов типифицированные названия.
Латрейль (Latreille 1802) впервые использовал в классификации членистоногих ранг семейства (familia), поставив его между родом и отрядом (это парадоксально, поскольку в латинском языке слово «genus» относится к более общей группе людей, чем слово «familia»); одновременно с этим он ввел дополнительные ранги – как выше семейства, так и ниже его. Исходно названия зоологических семейств были как типифицированными, так и нетипифицированными, а типифицированные не были унифицированы (см. ниже, I.6.2). Уже в первой половине XX века стало общепринятым образовывать названия таксонов группы семейства от названий типовых родов, добавляя к ним стандартные окончания, соответствующие определенным рангам (т.е. создавать типифицированные унифицированные названия). Таким образом появился удобный способ добавлять ранги, необходимые для классификации возрастающего числа новооткрытых видов, и этот способ относится только к рангам выше рода.
Для добавления рангов ниже рода удобного способа нет. В зоологии между родом и видом может быть добавлен только один ранг – подрод (в ботанике принято несколько внутриродовых рангов).
Использование ранга подрода имеет свои неудобства, из-за которых многие зоологи его избегают: подродовое название не отличается от родового, поэтому чтобы их различить, приходится указывать ранг; в биномене подродовое название пишется в скобках между родовым и видовым названиями, делая биномен более громоздким.
В результате этого, когда в каком-либо роде число известных видов возрастает настолько, что возникает необходимость ввести новый ранг, новый ранг вводят не внутри рода, а выше рода. Из-за этого происходит инфляция родов: по мере увеличения числа видов родовой ранг все время перемещается на подчиненный таксон, так что число родов быстро растет, а род все время оказывается самым мелким надвидовым таксоном. Так, если ранее был установлен род, удовлетворяющий всем требованиям систематики (голофилетический, с ясным диагнозом), то в дальнейшем его разделяют на несколько родов не потому, что возникли какие-либо сомнения в его естественности, а лишь потому, что увеличилось число различаемых видов.
Неудобства, связанные с неравноценностью рангов и инфляцией родов, устраняются, если использовать безранговый принцип систематики и безранговую номенклатуру (см. ниже, I.6.4, I.6.5, I.6.7 и главы III–X).