|
|
Международная научная
конференция
ФУНДАМЕНТАЛЬНЫЕ
ПРОБЛЕМЫ ЭНТОМОЛОГИИ
В XXI ВЕКЕ
Санкт-Петербург, 16–20 мая 2011 г.
Кладоэндезис
и новый взгляд на эволюцию метаморфоза у
насекомых
Cladoendesis
and a new
look at the evolution of insect metamorphosis
Клюге
Н.Ю.
Kluge
N.J.
Кафедра энтомологии Санкт-Петербургского государственного университета
Энтомологическое
обозрение, 2012
т.91, вып.1, стр.63–78 
английский
перевод:
Entomological Review, 2012
volume 92, issue 6, pp. 622–632
|
ENGLISH
|

эмблема
конференции, нарисованная автором
статьи; согласно принципам
кладоэндезиса, такого насекомого быть
не может
|
 |
|
 Введение
Введение
1. Принципы кладоэндезиса
2. Консервация признаков
3. Кладоэндезис и изучение метаморфоза
3.1. Amyocerata: эволюция линочных процессов в антенне
3.2. Metabola: полное превращение
3.3. Gallinsecta: иллюзия конвергенции
Литература
Summary
|
Кладоэндезис – это метод анализа филогении путем построении классификации,
в которой каждой филогенетической ветви
соответствует голофилетический таксон, а
каждой неразрешенной области
филогенетического древа соответствует
плезиоморфон, и все таксоны (голофилетические
и плезиоморфоны) снабжены собственными
названиями; характеристика каждого таксона
снабжена ссылками на характеристики вышестоящих таксонов, так что признаки всех таксонов формулируются как иерархически взаимосвязанные. |
Введение
В современной литературе "филогенетическим анализом" или "кладистическим анализом" зачастую называют вещи, заведомо не имеющие отношения к филогении (например, анализ внутривидовых форм), либо не соответствующие научному мировоззрению (например, анализ филогении на основе экономности эволюции, отвергаемой имеющимися теориями).
В последнее десятилетие мною был разработан эффективный метод анализа филогении, представляющий собой прямое продолжение и усовершенствование традиционного изучения филогении путем построении естественной классификации. Этот метод предложено называть
кладоэндезисом.
Изначально принципы кладоэндезиса были разработаны для изучения филогении поденок (Ephemeroptera)
(Kluge 2004a); они же позволили продвинуться в понимании гомологии гениталий насекомых (Клюге
2003), выявить ошибку в описании строения ног личинок скорпионниц – Mecoptera
(Kluge 2004b) и получить новые представления о метаморфозе насекомых. Благодаря применению принципов кладоэндезиса удалось существенно продвинуться в понимании того, что представляет собой
метаморфоз в различных таксонах насекомых, и как происходит линочная трансформация отдельных придатков
(Kluge 2005a ,
2010d); изучение процессов линочных трансформаций, помимо прочего, позволили выявить гомологию частей имагинальной максиллы у блох – Aphaniptera (Клюге
2002) и личиночной максиллы у двухоботных сетчатокрылых – Neuropteroidea-Birostrata
(Kluge 2005b).
Здесь мы обсудим некоторые моменты эволюции онтогенетической трансформации ног и антенн насекомых в свете последовательного применения принципов кладоэндезиса.
1. Принципы кладоэндезиса
Термин "кладоэндезис" предложен в недавно опубликованной статье (Kluge &
Novikova 2011) для обозначения того подхода к реконструкции филогении, который разрабатывается мною уже более десяти лет. Термин "кладоэндезис" (cladoendesis), то есть "ветвесцепление" (κλαδος + εν-δεσις) связан с тем, что при этом подходе особое внимание уделяется связи апоморфий каждого таксона с характеристиками вышестоящих таксонов, так что признаки всех таксонов изначально формулируются как иерархически взаимосвязанные.
В этом отношении кладоэндезис противопоставляется модным сейчас
матричным методам реконструкции филогении, при которых все признаки формулируются как независимые ни от чего и первоначально заносятся в прямоугольную матрицу, а не в иерархическую систему. При матричных методах филогения каждый раз реконструируется с нуля, как если бы до того ее никто никогда не реконструировал. Такой подход мог бы иметь смысл, если бы существовали методы анализа филогении, позволяющие получить окончательный результат, подобно тому, как это имеет место, например, при химическом анализе. Однако, поскольку при реконструкции филогении мы вынуждены выявлять эволюционные акты, однократно произошедшие в прошлом, здесь ни один метод анализа не может гарантировать правильный результат. Поэтому бессмысленно получать филогенетические реконструкции разными методами, чтобы потом сравнивать результаты. Единственным действенным способом реконструкции филогении является привлечение всех возможных фактов и теорий для построения одной единственной филогенетической реконструкции с тем, чтобы максимально приблизить ее к истинной филогении, понимая при этом, что получить окончательное знание филогении с заранее заданной точностью в принципе невозможно. Поэтому, даже если не принимать во внимание заведомую ненаучность такого популярного метода матричного анализа, как анализ парсимонии, следует понимать, что любые методы матричного анализа являются бесперспективными.
Кладоэндезис является прямым продолжением того традиционного метода анализа филогении, который был создан задолго до появления эволюционной теории и который породил саму эволюционную теорию и представление о филогении. Важнейшими вехами развития кладоэндезиса можно считать работы Аристотеля (который усмотрел
иерархическую
закономерность в разнообразии животных), К. Линнея (который разработал эффективный метод описания этой закономерности), французских авторов времен революции – Ж. Кювье, Ж.-Б. Ламарка, П.-А.
Латреиль и других (которые поставили вопрос о природе этой закономерности) и Ч. Дарвина (который, объяснив механизм эволюции, сделал эволюционный подход в биологии общепризнанным). Эти авторы и их последователи верили в то, что иерархическая упорядоченность разнообразия животных реально существует, и что если описать ее в виде иерархической классификации, то это позволит понять ее природу. Ч. Дарвин пришел к ясному выводу, что строившаяся со времен Линнея иерархическая классификация живых организмов отражает их генеалогию (т.е. филогению по терминологии Э. Геккеля):
"... Таким образом, мы имеем здесь много произошедших от общего предка видов, которые группируются в роды, роды – в подсемейства, семейства и отряды, а все вместе – в один большой класс. Так, по моему мнению, объясняется важный факт естественного распределения организмов в группы, подчиненные одна другой ..."
("... So that we here have many species descendent from a single progenitor grouped into genera ; and the genera in sub-families, families, and orders, all in one great class. Thus, the grand fact of the natural subordination of all organic beings in groups under groups ... is in my judgment explained.")
(Darwin 1871: 373). Однако после работ Дарвина в сознании некоторых биологов произошло раздвоение понятий: они считают, что классификация, которую начали создавать раньше, чем открыли филогению, не может быть отражением филогении, а представляет собой что-то иное (хотя что именно иное, никто из них объяснить не может). В действительности же
"никогда не было сделано попытки дать этому другое объяснение"
("no other explanation has ever been attempted")
(Darwin 1871: 373). Поэтому важным этапом в становлении кладоэндезиса являются работы В. Хеннига, который ввел понятие парафилии и ясно сформулировал принципы, по которым строится классификация, отражающая филогению
(Hennig 1950).
Главным принципом кладоэндезиса является то, что все данные записываются в виде классификации, в которой все признаки отнесены к иерархически соподчиненным таксонам. При этом новые данные вписываются в уже имеющуюся классификацию, которая по мере накопления новой информации дополняется и детализируется, а при выявлении ошибок – исправляется. Для того, чтобы выполнять эту функцию, классификация должна быть способной вмещать в себя все вновь добываемые факты и позволять легко находить их. Разработанные Линнеем
(Linnaeus 1758) и Латреилем (Latreille 1802) принципы классификации в значительной степени обеспечили решение этой задачи: благодаря им накопление данных достигло такого уровня, который позволил создать теорию эволюции и реконструировать ту часть филогении, которая нам сейчас известна. Однако нерешенной проблемой до недавнего времени оставалось то, что существовавшие принципы систематики и номенклатуры ограничивали классификацию заранее заданным набором искусственных абсолютных рангов, и поэтому не позволяли удобным способом записывать данные обо всех выявляемых иерархически соподчиненных филогенетических ветвях.
Для решения этой проблемы мною была разработана безранговая номенклатура, точнее, система из двух номенклатур
("dual nomenclature system"), которая, не противореча ни общепризнанному Международному кодексу зоологической номенклатуры, ни традициям в употреблении названий высших таксонов, позволяет снабдить однозначными названиями неограниченное число таксонов, распределенных по неограниченному числу иерархических уровней (Клюге
1999 ,
Kluge 1999 ,
Клюге 2000, Kluge
2009a, 2010с ). В отличие от умозрительных идей о модернизации номенклатуры, выдвигавшихся некоторыми авторами, данная номенклатура успешно используется на практике: в частности, благодаря ей оказалось возможным произвести полноценную
ревизию филогенетической системы поденок – Ephemeroptera
(Kluge 2004a, 2007a, 2007b, 2008, 2009b, 2010a, 2010b, Kluge & Novikova 2011).
Помимо рациональной номенклатуры, новой особенностью кладоэндезиса является рациональное изложение материала. Вместо повторяющихся разделов, таких как диагноз, описание, сравнение и др., каждый признак записывается один раз в характеристике того таксона, аутапоморфией которого этот признак является. При этом в описании признака даются ссылки на признаки вышестоящих таксонов, относящихся к той же детали строения; таким образом аутапоморфии каждого таксона в явном виде сравниваются со строением, исходным для вышестоящих таксонов.
Исходя из предположения, что каждый признак, обладающий эволюционным консерватизмом (см. 2) и таксономической значимостью, является аутапоморфией некоего голофилетического таксона, можно выявлять новые голофилетические таксоны и, таким образом, продвигаться в реконструкции филогении.
Если классификация и номенклатура таксонов ограничиваются заранее определенным набором рангов, создание новых голофилетических таксонов может оказаться невозможным; безранговая же номенклатура позволяет использовать классификацию для реконструкции филогении без каких-либо ограничений.
Важное значение для кладоэндезиса имеет введение термина "плезиоморфон"
(Kluge 2004a: 15 ). Плезиоморфон – это таксон, характеризующийся только плезиоморфиями, так что нет оснований считать этот таксон голофилетическим; в то же время парафилия этого таксона также может быть не доказанной. Для доказательства парафилии таксона необходимо найти синапоморфию между частью этого таксона и другим таксоном; в этом случае появляется возможность расформировать прежний парафилетический таксон, объединив его часть с другим таксоном на основании найденной апоморфии, которая в этом случае становятся аутапоморфией нового таксона. Таким образом, в филогенетической классификации нет заведомо парафилетических таксонов: как только парафилия таксона становится известной, таксон расформировывают. Однако в любых классификациях неизбежно присутствуют плезиоморфоны: их парафилия лишь предполагается, но отсутствие прямого доказательства парафилии (т.е. отсутствие знаний о соответствующих апоморфиях) не позволяет расформировать эти таксоны. Необходимо ясно различать голофилетические таксоны и плезиоморфоны. Для этого предлагается перед названием каждого плезиоморфона писать слово "плезиоморфон" (или латинское "plesiomorphon"), а в таблицах, схемах и указателях, где требуется сокращение, писать сокращенное "pm" или "pm." – например, "plesiomorphon Permoplectoptera", или "pm.Permoplectoptera". Употребление похожего термина "плезион" вместо термина "плезиоморфон" является ошибочным
(Kluge 2004a ).
2. Консервация признаков
Прежде, чем перейти к обсуждению эволюции метаморфоза, следует сказать о важном эволюционном механизме, не имеющем пока ясного объяснения – а именно о консервации признаков. В ходе эволюции признаки не только возникают и исчезают, но и консервируются и деконсервируются. Консервация признака выражается в том, что он становится неспособным к дальнейшим эволюционным изменениям. Если этот признак оказывается вредным, эволюция не может его уничтожить, но может тем или иным способом привести к тому, что этот признак перестает мешать выживанию. Поэтому стабильность такого признака никак не может быть объяснена действием естественного отбора; в ее основе должен лежать какой-то генный механизм. Консервация признаков не поддается изучению методами экспериментальной генетики, поскольку признак, оказавшийся законсервированным, не может быть изменен в эксперименте. Явление консервации признаков хорошо известно в традиционной систематике: таксономисты выискивают законсервированные признаки, чтобы использовать их в качестве диагностических признаков таксонов. Большая часть апоморфий, на которых основаны известные нам представления о филогении, являются законсервированными: в отличие незаконсервированных апоморфий, их можно проследить во всем голофилетическом таксоне и тем самым убедиться в его голофилии.
Бытует мнение, будто существуют какие-то более консервативные и менее консервативные категории признаков (например, морфологические признаки якобы консервативнее поведенческих, мускулатура якобы консервативнее скелета и т.п.). Некоторые систематики пытаются убедит себя в том, что если признак оказался таксономически значимым в одном таксоне, то подобный ему признак, найденный в другом таксоне, будет иметь такое же таксономическое значение. Это было бы удобно для практической работы систематиков, однако это не так. В действительности, как совершенно правильно писал Карл Линней
"то, что в одном роде важно для установления рода, в другом – вообще не имеет значения"
("Quae in uno genere ad Genus stabiliendum valent, minime idem in altero necessario praestant")
(Linnaeus 1751: 114, §169). Каждый законсервированный признак является конкретным признаком конкретного таксона, и его консервация является однократным эволюционным актом. Из-за того, что законсервированными и таксономически значимыми являются конкретные признаки конкретных таксонов, а не совокупности аналогичных признаков, таксономические признаки не могут быть адекватно записаны в матрицу таксон/признак, что делает бессмысленными все матричные методы филогенетического анализа.
В кладоэндезисе представление о законсервированных признаках играет ключевую роль, поскольку традиционно строящаяся естественная классификация основана главным образом на выявлении законсервированных признаков среди прочих.
В качестве примера законсервированного признака можно привести наличие шести ног у Hexapoda – точнее, наличие трех сегментов груди, каждый из которых в принципе может иметь по одной паре ног. При всем огромном разнообразии насекомых далеко не всем им нужно иметь три пары ног: если ноги оказываются лишними, они не функционируют (как, например, передние ноги Nymphalidae) или утрачиваются; если трех пар ног оказывается недостаточным, появляются дополнительные локомоторные органы (например, ложноножки). Но общее число сегментов, дифференцированных в качестве грудных, неизменно остается равным трем. В экспериментах на дрозофилах удается изменить строение того или иного грудного сегмента (например, при мутациях bithorax заднегрудь становится сходной со среднегрудью и несет такую же пару крыльев), но общее число сегментов груди изменить не удается. У других членистоногих аналогичный признак (число сегментов груди или аналогичной ей тагме) может не иметь такой консервативности. Например, у морских пауков (Pantopoda) имеется 4 сегмента с ходильными ногами, но у отдельных видов их число увеличено до 5 или 6. У ракообразных (Eucrustacea) число сегментов груди различается в разных таксонах, а в некоторых случаях (например, у Notostraca и некоторых Anostraca) варьирует индивидуально.
Представление о консервировании признаков очень важно для понимания эволюции метаморфоза.
3. Кладоэндезис и изучение метаморфоза
Эволюцию метаморфоза насекомых зачастую рассматривают вне связи с филогенией; некоторые авторы предлагали классификации типов метаморфоза, отличные от филогенетической классификации насекомых. В действительности каждый тип метаморфоза возник однократно и присущ единственной филогенетической ветви.
Филогенетическая классификация обсуждаемых здесь таксонов представляется следующим образом
 :
:
1. Hexapoda Blainville 1816 (nom.hier.: Scarabaeus/fg3)
1.1. Entognatha Stummer-Traunfels 1891 (nom.hier.: Podura/fg1)
1.2. Amyocerata Remington 1955 (nom.hier.: Scarabaeus/fg4)
1.2.1. Triplura Ewing 1942 (nom.hier.: Lepisma/fg1)
1.2.2. Pterygota Gegenbaur1878 (nom.hier.: Scarabaeus/fg5)
1.2.2.1. Ephemeroptera Hyatt & Arms 189 (nom.hier.: Ephemera/fg1)
1.2.2.2. Metapterygota Börner 1909 (nom.hier.: Scarabaeus/fg6)
1.2.2.2.1. Odonata Fabricius 1793 (nom.hier.: Libellula/fg1)
1.2.2.2.2. Neoptera Martynov 1923 (nom.hier.: Scarabaeus/fg7)
1.2.2.2.2.1. Plesiomorphon Polyneoptera Martynov 1923 (nom.hier.: Gryllus/f1=Forficula/g1)
1.2.2.2.2.2. Eumetabola Hennig 1953 (nom.hier.: Scarabaeus/fg8)
1.2.2.2.2.2.1. Metabola Burmeister 1832 (nom.hier.: Scarabaeus/fg9)
1.2.2.2.2.2.2. Parametabola Crampton 1938 (nom.hier.: Cimex/f1=Cicada/g1)
1.2.2.2.2.2.2.1. Zoraptera Silvestri 1913 (nom.hier.: Zorotypus/fg1)
1.2.2.2.2.2.2.2. Acercaria Börner 1904 (nom.hier.: Cimex/f2=Cicada/g2)
1.2.2.2.2.2.2.2.1. Panpsocoptera Crampton 1938 (nom.hier.: Psocus/f1=Pediculus/g1)
1.2.2.2.2.2.2.2.2. Condylognatha Börner 1904 (nom.hier.: Cimex/f3=Cicada/g3)
1.2.2.2.2.2.2.2.2.1. Thysanoptera Haliday 1836 (nom.hier.: Thrips/fg1)
1.2.2.2.2.2.2.2.2.2. Arthroidignatha Spinola 1850 (nom.hier.: Cimex/f4=Cicada/g4)
1.2.2.2.2.2.2.2.2.2.1. Hemelytrata Fallen 1829 (nom.hier.: Cimex/f5=Cicada/g5)
1.2.2.2.2.2.2.2.2.2.2. Plantisuga Dumeril 1806 (nom.hier.: Aphis/fg1)
1.2.2.2.2.2.2.2.2.2.2.1. Psyllaleyroda Kluge 2010 (nom.hier.: Psylla/fg1)
1.2.2.2.2.2.2.2.2.2.2.1.1. Saltipedes Amyot & Serville 1843 (nom.hier.:
Psylla/fg2)
1.2.2.2.2.2.2.2.2.2.2.1.2. Scytinelytra Amyot & Serville 1843 (nom.hier.:
Aleyrodes/fg1)
1.2.2.2.2.2.2.2.2.2.2.2. Aphidococca Kluge 2010 (nom.hier.: Aphis/fg2)
1.2.2.2.2.2.2.2.2.2.2.2.1. Gynaptera Laporte 1834 (nom.hier.: Aphis/fg3)
1.2.2.2.2.2.2.2.2.2.2.2.2. Gallinsecta De Geer 1776 (nom.hier.: Coccus/fg1)
Здесь аббревиатура "nom.hier." означает типифицированное безранговое иерархическое название (nomen hiererchicum) того же таксона, для которого приведено нетипифицированное безранговое циркумскриптное название (nomen circumscribens). Обсуждения статусов названий, используемых в данной классификации, даны в предыдущих статьях
(Kluge 2010c, 2010d).
При описании метаморфоза, наряду с общепринятыми терминами "личинка первого возраста", "личинка второго возраста", "нимфа второго возраста" и т.п. и соответствующих им сокращений "L1", "L2", "N2", оказалось необходимым использовать латинские термины
(Kluge 2010d). Каждый такой термин представляет собой одно сложное слово –
примоларва (primolarva), секундоларва (secundolarva),
секундонимфа (secundonympha) и т.п. При описании процессов трансформации при линьке с одного возраста на другой используются прилагательные, образованные от этих терминов ("примоларвалный", "секундоларвалный" и т.п.), что было бы невозможным при многословном обозначении возрастов. Старинные термины "личинка" и "нимфа" не имеют общепринятых определений, поэтому при назывании возрастов следует везде использовать сквозную нумерацию: например, если первые два возраста считаются личиночными, а следующие нимфальными, то после секундоларвы следует тертионимфа (а не примонимфа). Поскольку последние возраста бывают более стабильными, чем предшествующие, для их обозначения можно использовать термины
ультимонимфа (ultimonympha) и пенультимонимфа (penultimonympha), или соответствующие им
ультимоларва (ultimolarva) и пенультимоларва (penultimolarva). Для соблюдения единой сквозной нумерации возрастов у насекомых с полным превращением личинку последнего возраста следует называть "пенультимоларва"; в данном случае термины "ультимоларва" или "ультимонимфа" не используются, поскольку последний преимагинальный возраст имеет общепринятое название "куколка" (pupa).
Некоторые авторы, описывая метаморфоз насекомых с полным превращением, утверждали, что в ходе его происходит повторная закладка конечностей или даже повторный эмбриогенез. Имагинальные диски круглошовных мух (Cyclorapha) иногда описывают как некие "эмбриональные" ткани, скрывшиеся внутри тела личинки, чтобы заново пройти что-то вроде эмбриогенеза и породить взрослую муху. Принципы кладоэндезиса не допускают подобных фантазий, поскольку требуют рассматривать метаморфоз любого организма в контексте уже установленного филогенетического положения этого таксона. Насекомые, о которых идет речь, относятся к
Hexapoda
Blainville 1816, которые относятся к
Arthropoda Siebold 1848, которые, в свою очередь, относятся к
Monostomata
Huxley 1875 (=
Eumetazoa Butschli 1910). Таксон Monostomata, помимо прочего, характеризуется тем, что на ранних этапах эмбриогенеза происходит необратимая дифференциация бластодермы на эктодерму и энтодермально-мезодермальный зачаток. Arthropoda, помимо прочего, характеризуются однократной закладкой конечностей каждого сегмента. Hexapoda, помимо прочего, характеризуются уникальным и очень консервативным планом строения грудных конечностей (ног), не встречающимся ни у каких других членистоногих. В то же время для всех Arthropoda свойственна способность к регенерации конечностей: при ампутации части конечности из оставшейся части развивается нормальная конечность со всеми свойственными ей члениками (например, у Hexapoda из уцелевшего тазика с вертлугом может развиться нога, состоящая из тазика, вертлуга, бедра, голени и лапки). Таким образом, при регенерации нарушается онтогенетическая гомология члеников конечности, но сохраняется онтогенетическая гомология конечности в целом (о термине "онтогенетическая гомология" – см.
Kluge 2005a ). Эти же возможности реализуются и в нормальном развитии при любых модификациях метаморфоза: в случаях замены конечностей новая конечность возникает не заново и не из какого-то мифического "имагинального диска", а развивается из остатков тканей предшествующей конечности; расчленение же конечности может при этом возникать совершенно заново. Даже такие причудливые образования, как имагинальные диски Cyclorapha, представляют собой лишь особым образом модифицированные предлиночные складки покровов, свойственные всем членистоногим
(Kluge 2005a).
3.1. Amyocerata: эволюция линочных процессов в антенне
Принято считать, что при наиболее примитивном метаморфозе развитие идет наиболее прямым путем, а такие явления, как замена органа путем его отмирания и последующего восстановления, являются вторичными. Однако, это не всегда так.
Таксон Amyocerata Remington 1955
часто неверно называют "Ectognatha"
или "Insecta", что порождает путаницу и заблуждения (Клюге
1996, Kluge 1999, Клюге 2000, Kluge 2010c). В действительности Amyocerata – это голофилетический таксон в составе голофилетического таксона Hexapoda. Наиболее яркой аутапоморфией Amyocerata является строение антенн: исходно антенна состоит из одночленикового скапуса, одночленикового педицеллюса и многочленикового флагеллума; к основанию скапуса подходят мышцы, идущие от тенториума или иной скелетной опоры внутри головы; скапус вмещает мышцы, идущие от его основания к основанию педицеллюса; других мышц в антенне нет; в ходе эволюции антенн у амиоцерат может происходить лишь редукция мышц, но не появление новых мышечных члеников. Следует обратить внимание на очень высокую консервативность этих признаков: независимо от размера, формы и функции антенн у амиоцерат, у них никогда не бывает мышц в иных члениках кроме скапуса; в то же время мышцы скапуса устойчиво сохраняются даже при сильном укорочении антенны, и редуцируются лишь в исключительных случаях. С тех пор как этот признак был выявлен Иммсом
(Imms 1939) не было обнаружено никаких исключений.
В ходе онтогенеза при каждой линьке скапус (с находящимися в нем мышцами) и педицеллюс сохраняются, меняя лишь размер и/или форму, а флагеллум обычно либо также сохраняет прежнее число члеников, либо число его члеников увеличивается. В случаях увеличения числа члеников флагеллума, это достигается путем роста и деления наиболее проксимального членика; иногда имеется также одна или несколько дополнительных зон роста в других участках флагеллума
(Imms 1940). Интересно, что в ряде случаев, одновременно с добавлением некоторого числа члеников в проксимальной части флагеллума, происходит отмирание и отбрасывание некоторого числа дистальных члеников флагеллума; в ходе такой линьки общая длина флагеллума и общее число его члеников обычно увеличиваются. Остатки тканей отмерших члеников флагеллума не рассасываются полностью, а сбрасываются вместе с линочной шкуркой, так что такой тип линьки может быть выявлен при изучении пустых линочных экзувиев (в отличие от сходных процессов в конечностях у некоторых насекомых, при которых все остатки рассасываются). У антенны, линяющей таким способом, вершинный членик не имеет каких-либо морфологических особенностей, поскольку при каждой линьке функция вершинного членика переходит к одному из члеников, бывшего до этого срединным. В некоторых случаях отбрасывание дистальной части флагеллума можно было бы объяснить механическими причинами: вершинная часть флагеллума бывает такой тонкой, что при экдизисе находящиеся в ней ткани не могут выйти из старой кутикулы и обрываются. Однако в некоторых случаях обрыв тканей происходит в таком месте, где толщина флагеллума не препятствует линьке, и этот обрыв образуется не при экдизисе, а формируется задолго до него (Kluge 2010d: Fig.
24).
Линьки с таким предопределенным обрывом дистальной части флагеллума встречаются у Triplura (как у Zygentoma, так и у Microcoryphia), у Ephemeroptera и у Plecoptera. Судя по систематическому положению этих таксонов, такой парадоксальный способ роста флагеллума является исходным для Amyocerata.
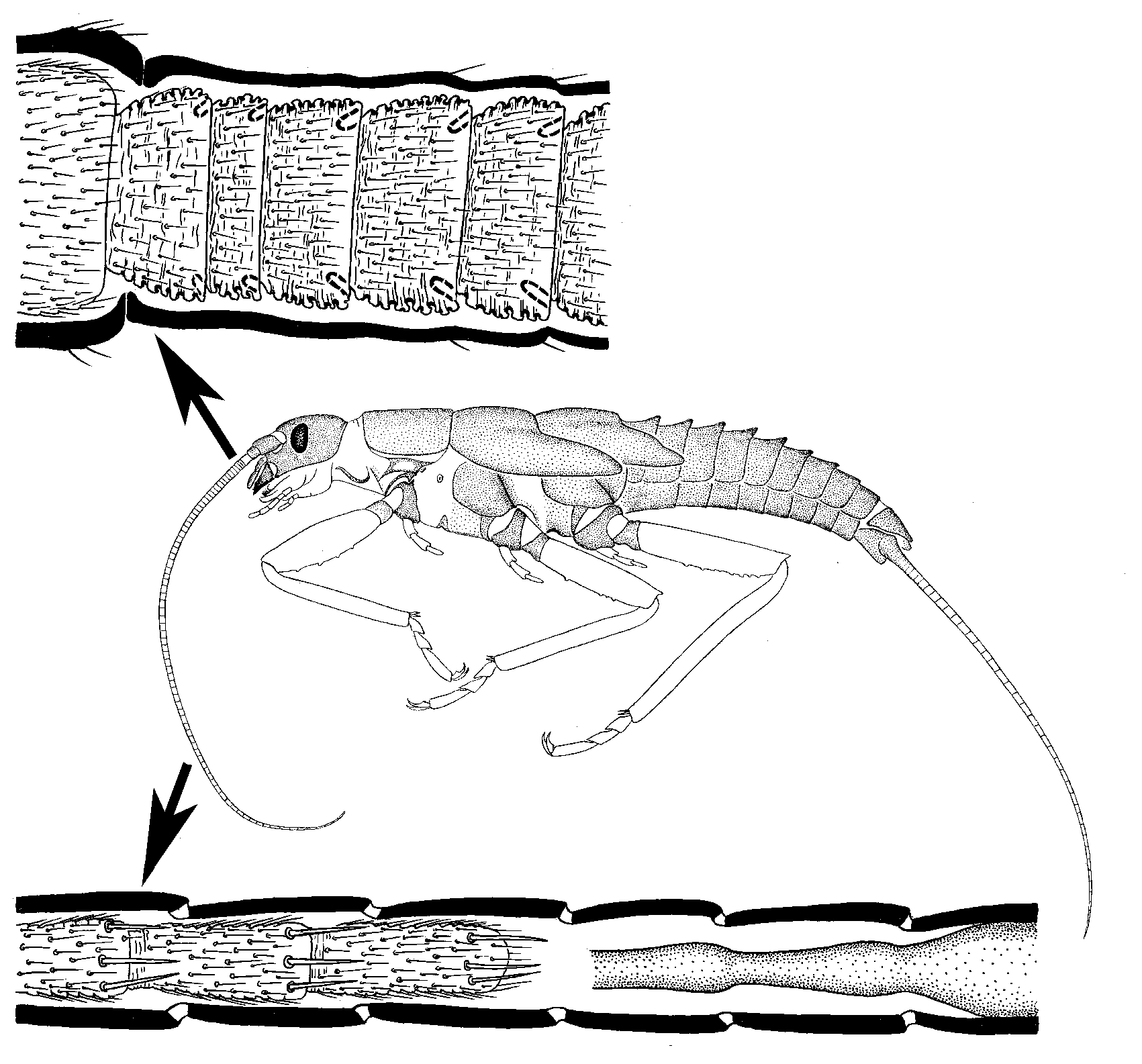
Подобный процесс добавления проксимальных члеников и отмирания дистальных члеников происходит и у многочлениковых каудалий Triplura, Ephemeroptera и Plecoptera.
У поденок (Ephemeroptera) в ходе предпоследней линьки многочлениковый флагеллум личинки превращается в тонкий нерасчлененный кольчатый флагеллумом субимаго. Как и при предшествующих личиночных линьках, дистальная часть живой ткани флагеллума отмирает, обрывается и остается в личиночном экзувии. Это происходит независимо от того, уменьшается ли при этой линьке общая длина флагеллума, или нет.
У Odonata, многих Polyneoptera (Embioptera, Isoptera, Dermatoptera, Spectra, и др.) и у всех Parametabola флагеллум антенны имеет постоянную вершину, не отбрасываемую при линьках. Судя по всему, такой способ развития возник независимо в разных филогенетических ветвях. У Cercopoidea (относящихся к Parametabola) при последней линьке происходит преобразование флагеллума, внешне сходное с личиночно-субимагинальным преобразованием флагеллума у поденок: 7-члениковый флагеллум личинки превращается в нерасчлененный кольчатый флагеллум имаго; но при этом, в отличие от поденок, никакие ткани не отбрасываются. В таксоне Gallinsecta (также относящемся к Parametabola) возникло совершенно необычное линочное преобразование всей антенны, при котором дистальная часть ткани флагеллума отмирает и отбрасывается (см. 3.3); несмотря на это, антенна может иметь оформленные апикальные структуры, всегда находящиеся только на последнем членике
(Kluge 2010d).
У всех насекомых с полным превращением (Metabola) антенна личинки утратила строение, свойственное Amyocerata (см. 3.2). Несмотря на глубокие преобразования, происходящие при линьке с личинки на куколку и далее на имаго, никакого отмирания и отбрасывания тканей в антенне не происходит. Принято считать, что полное превращение отличается от других типов метаморфоза более глубокими некротическими процессами; однако в отношении вершины антенны дело обстоит как раз наоборот: исходно для Amyocerata она развивается с некрозом, а у Metabola это свойство утрачено.
3.2. Metabola: полное превращение
Несмотря на общеизвестность полного превращения и обилия литературы о нем, до последнего времени было неясно, в чем именно оно выражается. Из поверхностных наблюдений видно, что в цикле развития насекомых с полным превращением есть (1) подвижная питающаяся стадия личинки, чем-то напоминающая червя, (2) неактивная стадия куколки, способная совершать лишь ограниченные движения или совсем неподвижная, и (3) стадия имаго. При этом оставалось неясным, что именно вызывает у большинства людей ассоциацию личинок с червями (при том, что строение личинок очень разнообразно) и в чем именно ограничена подвижность у всех куколок.
Уже к началу XX века стало общепризнано, что все насекомые с полным превращением образуют голофилетический таксон Metabola. Путаницу в это утверждение вносит лишь тот факт, что вместо правильного названия
Metabola
Burmeister 1832 зачастую используются неправильные названия "Holometabola" и "Endopterygota", тогда как исходно названия
Holometabola
Burmeister 1835 и
Endopterygota Sharp 1899 были введены для таксонов иного состава. В современной литературе для этого таксона часто используют название Oligoneoptera
Martynov 1923, являющееся младшим циркумскриптным синонимом названия Metabola (Клюге2000,
Kluge 2010c).
Доказательства голофилии Metabola можно сформулировать в виде следующего утверждения: в любом таксоне, голофилия которого надежно установлена на основании бесспорных аутапоморфий
1) либо все без исключения виды имеют полное превращение,
2) либо все без исключения виды имеют метаморфоз, отличный от полного превращения,
3) либо (если это больший таксон, включающий в себя Metabola) включены все без исключения виды, имеющие полное превращение.
Проверить это утверждение прямым путем невозможно, поскольку все виды насекомых еще неизвестны, а среди тех, что формально описаны, большая часть известна лишь по отдельным взрослым экземплярам. В XIX веке метаморфоз был известен для такого малого числа видов, что правильность этого утверждения могла вызывать сомнение. К настоящему времени число видов, для которых метаморфоз известен, столь велико, что находка какого-либо исключения из сформулированного выше утверждения представляется невероятной. Сегодня еще можно предположить находку неизвестного вида бабочек с необычными морфологическими признаками, но уже невозможно предположить, что будет обнаружена бабочка, не имеющая полного превращения. Утверждение о голофилии Metabola полностью основывается только на принципах кладоэндезиса: оно вытекает из огромного опыта неопределенно большого числа систематиков и других исследователей и наблюдателей, обобщенного в обширной таксономической литературе, не поддающейся формальному анализу. До настоящего времени исследователи даже не знали, чем собственно характеризуется полное превращение, однако это не мешало им правильно узнавать этот тип метаморфоза и отличать его от других.
Многие авторы пытались объяснить происхождение полного превращения какими-то причинами – экологическими, морфогенетическими, эмбриологическими, либо иными. Однако, если Metabola является голофилетисческим таксоном, никакие из этих объяснений не могут быть верными: если бы существовала какая-нибудь причина, приведшая к появлению полного превращения, она приводила бы к повторному появлению такого же типа метаморфоза, и в таком случае таксон, объединяющий всех насекомых с полным превращением, не был бы голофилетическим. Голофилия Metabola означает, что полное превращение возникло у одного позднепалеозойского вида, который существовал одновременно с множеством других видов крылатых насекомых; этот эволюционный акт был случайным в такой степени, что ни разу больше не повторился. Таким образом, следует ставить вопрос не о том, почему возникло полное превращение, а о том, какие именно конкретные свойства возникли у общего предка Metabola и были унаследованы всеми его потомками. С этой точки зрения полное превращение почти никто не рассматривал. В моей статье
(Kluge 2005a ) сделана первая попытка сформулировать конкретные аутапоморфии Metabola, выявленные при изучении лишь нескольких представителей.
Ключевой особенностью Metabola оказалась своеобразная трансформация ног при линьке с личинки на
куколку: живые ткани ноги обычно сначала дегенерируют, сильно уменьшаясь в размерах и утрачивая мышцы, а затем растут так, что всегда образуется неподвижный коленный сгиб; поэтому после сбрасывания личиночной кутикулы у куколка ноги всегда оказываются согнутыми в коленях и лишенными мышц, которые могли бы их разогнуть. Эта частная особенность законсервирована настолько прочно, что никакие эволюционные изменения не были способны ее изменить. В результате этого у всех Metabola куколки теми или иными способами приспособились к существованию с неподвижными ногами: в большинстве случаев это неактивное нахождение в укрытии, в более редких случаях (напимер, у Culicomorpha) это приспособление к такому движению, при котором неподвижные ноги не мешают. Такими образом стало понятно, чем определяется известное свойство куколок делать бесполезные движения туловищем, не имея способности передвигаться.
В предыдущей статье (Kluge 2005a: 217) я, не имея данных по метаморфозу Hymenoptera-Symphyta, процитировал статью Ивановой-Казас и Ивановой (1964), где утверждается, что "никаких дегенеративных изменений в гиподерме ног при этом не наблюдаются"; на этом основании я отнес личинок Hymenopter-Symphyla к группе "коротконогих", вместе с Lepidoptera. Однако, как показали мои более поздние наблюдения за несколькими видами пилильщиков – Tenthredinoidea
(Diprion pini L., Cimbex femorata L. и др.), их ноги развиваются по типу ног "длинноногих" личинок, т.е. личиночно-куколочная трансформация ног сопровождается не только полной утратой мышц, но очень сильным уменьшением гиподермы.
Червеобразность большинства личинок Metabola, выражающаяся у разных представителей весьма по-разному, по-видимому объясняется тем, что личинка общего предка Metabola имела очень короткие ноги, сохранявшие полный набор члеников и мышц, свойственных Hexapoda (среди современных насекомых такие ноги присущи личинкам многих Lepidoptera); эта личинка передвигалась в значительной степени за счет движений туловища, которое имело червеобразную форму. Дальнейшая эволюция форм личинок шла в самых разных направлениях – и в сторону редукции ног до их полной утраты, и в сторону вторичного удлинения ног; поскольку эволюция личинок во всех случаях стартовала от червеобразного строения, большая часть личинок сохраняет те или иные черты этой исходной червеобразности. Однообразие роста куколочной ноги (с неизменным образованием коленного сгиба) и удивительное разнообразие способов предшествующей дегенерации части личиночной ткани ноги свидетельствует о том, что укорочение личиночных ног произошло однократно у исходного предка Metabola, а вторичное удлинение личиночных ног происходило многократно в разных таксонах в составе Metabola.
Новые наблюдения позволяют предположить, что все Coleoptera-Polyphaga имеют единый способ дегенерации личиночных ног с нарушением онтогенетической гомологии члеников, как это подробно описано мною для мучного хрущака
(Tenebrio molitor L.) и колорадского жука (Leptinotarsa decimlineata
(Say 1824)) (Kluge 2005a ). У Coleoptera-Adephaga способ дегенерации ног совершенно иной. У плавунца
(Dytiscus marginalis L.) и некоторых видов жужелиц (Carabidae) наблюдалось отмирание и растворение тканей дистальной части личиночной ноги без отслоения гиподермы от кутикулы, после чего куколочная нога развивается из тканей, оставшихся в проксимальной части личиночной ноги.
Помимо особой трансформации ног, таксон Metabola характеризуется уникальным развитием антенн: антенна личинки не имеет амиоцератного плана строения, у нее нет членика, соответствующего скапусу и нет никаких мышц внутри; если она многочлениковая, ни один ее членик не соответствует какому-либо членику имаго. При метаморфозе антенна имаго развивается из всей антенны личинки без отмирания какой-либо ее части, но полностью утрачивает личиночное расчленение и приобретает новое, имагинальное – т.е. антенна личинки и имаго в целом и онтогенетически, и филогенетически гомологичны, но их отдельные членики не имеют ни онтогенетической, ни филогенетической гомологии. Мышцы скапуса воникают лишь в процессе куколочного развития. В отличие от ног, которые у некоторых Metabola в конце куколочного развития могут приобретать подвижность, антенны никогда не приобретают подвижности до самой линьки на имаго.
Другие аутапоморфии Metabola описаны в той же статье
(Kluge 2005a ).
3.3. Gallinsecta: иллюзия конвергенции
Принято считать, что у кокцид (т.е. у червецов и щитовок –
Gallinsecta De
Geer 1776) метаморфоз самцов имеет сходство с полным превращением. Действительно, в ходе онтогенеза самец кокциды проходит стадию личинки, которая активно питается, не имеет протоптеронов и сильно отличается от взрослого насекомого; стадию нимфы, непитающейся и ведущей неподвижный образ жизни в укрытии; и стадию крылатого имаго, сильно отличающегося от личики. Сходство ультимонимфы Gallinsecta с куколкой Metabola усугубляется тем, что ее антенны неподвижно направлены назад
(Kluge 2010d: Fig. 4). У некоторых Metabola (Trichoptera, Rhaphidioptera, Chrysopidae и др.) куколка перед самой линькой на имаго (т.е. в состоянии фаратного имаго) приобретает способность разгибать ноги и ходить на них, но антенны при этом остаются неподвижно направленными назад
(Kluge 2005a: Fig.39).
Некоторые авторы даже утверждали, что метаморфоз кокцид представляет собой переходную форму между полным и неполным превращением, поскольку самки развиваются с неполным превращением, а самцы – с полным. Большинство авторов используют более обтекаемые выражения, говоря о конвергентном сходстве между метаморфозом самцов кокцид и полным превращением Metabola. Поскольку возникновение конвергенций никак не объясняется современной эволюционной теорией, термин "конвергенция" означает лишь сходство, существование которого мы не можем объяснить.
Червецы и щитовки образуют бесспорно голофилетический таксон Gallinsecta. Как видно из приведенной выше филогенетической классификации насекомых, у Gallinsecta и Metabola не может быть никаких общих признаков, которые отсутствовали бы в других таксонах Eumetabola. В данной классификации лишь таксон Parametabola имеет спорный статус (поскольку некоторые авторы считают, что Zoraptera не являются сестринской группой для Acercaria). Все прочие таксоны, разделяющие Metabola и Gallinsecta, являются бесспорно голофилетическими – это Acercaria, Condylognatha, Arthroidignatha, Plantisuga и Aphidococca. Здесь важно отметить, что для доказательства голофилии этих таксонов не понадобились и никогда не понадобятся никакие матричные методы анализа филогении: каждый из этих таксонов характеризуется уникальными аутапоморфиями, не встречающимися нигде более в живой природе (т.е. внешней группой является не произвольно выбранный вид, а совокупность всех живых организмов), и наличие этих аутапоморфий проверено неопределенно большим числом исследователей для всех известных им видов, что не поддается занесению ни в какую матрицу.
Необходимо было сравнить метаморфоз у самцов Gallinsecta с полным превращением у Metabola, чтобы выяснить, есть ли между ними конкретные черты сходства. До недавнего времени этого сделать было невозможно, поскольку было неясно, чем конкретно характеризуется полное превращение. Теперь, когда стало ясно, что ключевой особенностью полного превращения является особая трансформация ног, и что неподвижность антенн у куколок Metabola вызвана их особым строением у личинки, появилась возможность сравнить эти онтогенетические процессы у Metabola и Gallinsecta.
В качестве представителя Gallinsecta мною был исследован крапивный червец –
Orthezia urticae L. Трансформация ног и антенн при всех линьках обоих полов у этого вида подробно описана в предыдущей статье
(Kluge 2010d). Как выяснилось в ходе этого исследования, при линьках непитающихся "куколкоподобных" нимф самца не происходит никаких процессов, аналогичных трансформации ног и/или антенн при полном превращении: никакие их части не дегенерируют, все мышцы сохраняются, каждый членик следующего возраста развивается из соответствующего членика предыдущего возраста и перед линькой помещается внутри его кутикулы, так что на всех фазах линочного цикла все членики в принципе сохраняют способность к активному движению.
Тот факт, что пенультимонимфа и ультимонимфа все время неподвижно сидят в укрытии, по-видимому объясняется лишь тем, что им нет надобности передвигаться. У всех Gallinsecta самцы имаго не питаются, но способны летать и спариваться, так что при превращении личинки в имаго исчезает ротовой аппарат и появляются крылья и гениталии. На стадиях пенультимонимфы и ультимонимфы ротовой аппарат уже полностью исчез, а крылья и гениталии еще имеют вид неподвижных протоптеронов и протопениса, неспособных функционировать.
В ходе этого же исследования было обнаружено, что при линьках питающихся стадий (это два первых возраста самца и все возраста самки) происходит совершенно уникальная трансформация всех придатков – и ног, и антенн. При каждой такой линьке дистальная часть каждого придатка полностью отмирает, а оставшаяся проксимальная часть утрачивает расчленение и всю внутреннюю мускулатуру. Далее придаток растет, заново приобретая мускулатуру и расчленение и достигая размера больше прежнего; при этом росте первый членик придатка (соответственно, скапус антенны и тазик ноги) растет, впячиваясь в тело в вывернутом наизнанку состоянии, т.е. покровами внутрь и мышцами наружу. В таком состоянии эти членики пребывают до самого экдизиса, так что их вворачивание и приведение в нормальное положение (мышцами внутрь) происходит благодаря усилиям насекомого, когда оно вылезает из треснувшей старой кутикулы. У
O. urticae питающиеся стадии держатся на растении при помощи ног; такое функционирование ног на протяжении всего линочного цикла, при котором ноги претерпевают столь катастрофическую перестройку, возможно благодаря тому, что мышцы, идущие из туловища к проксимальным краям тазика, не дегенерируют и не меняют своего места прикрепления. В результате каждой такой линьки все членики ног и все членики антенн оказываются онтогенетически негомологичными члеником соответствующего придатка предыдущего возраста. У
O. urticae это не связано ни с какими морфологическими изменениями, поскольку внешне все следующие друг за другом возраста выглядят так, как будто они развивались поступательно, без какой-либо дегенерации и замены.
Судя по фрагментарным данным, имеющимся для нескольких других видов Gallinsecta, можно предположить, что такое парадоксальное развитие у
O. urticae является не исходным для Gallinsecta, а производным от более сложного развития, при котором после подвижной личинки первого возраста следует безногая личинка второго возраста, а далее – снова подвижная стадия с ногами и антеннами.
В любом случае метаморфоз Gallinsecta является уникальной аутапоморфией этого таксона: выворачивание части конечности наизнанку неизвестно ни у каких других насекомых, включая насекомых с такими необычными модификациями метаморфоза, как у белокрылок (Scytinelytra) или мух (Cyclorapha).
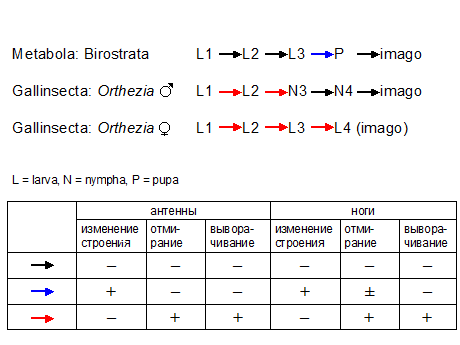
На таблице приведена упрощенная схема сравнения метаморфозов у самцов Gallinsecta и у Metabola, в которой в качестве представителя Gallinsecta взят крапивный червец
(Orthezia urticae), а в качестве представителя Metabola – златоглазка
(Chrysoperla carnea), поскольку у них совпадает число возрастов. Если составить такую схему, которая учитывает все разнообразие метаморфозов у Gallinsecta и у Metabola, она будет сложнее, но ее смысл будет тот же: модифицированные линьки происходят у Gallinsecta и у Metabola на разных стадиях и различаются по характеру модификаций.
Литература
Иванова-Казас О.М., Иванова Н.А. 1964. Метаморфоз ивового пилильщика
Pontania capreae L. (Hymenoptera, Tenthredinidae). I. Гиподерма // Энтомологическое обозрение
43 (2): 309–326.
Клюге Н.Ю. 1996. Мифы в систематике насекомых и принципы зоологической номенклатуры. // Энтомологическое обозрение
75 (4): 939–944.
Клюге Н.Ю. 1999. Система альтернативных номенклатур надвидовых таксонов. // Энтомологическое обозрение
78 (1): 224–243. Английский перевод: Kluge N.J. A system of alternative nomenclatures of supra-species taxa. Linnaean and post-Linnaean principles of systematics. // Entomological
Review 79 (2): 133–147.
Клюге Н.Ю. 2000. Современная систематика насекомых. Часть I. Принципы систематики живых организмов и общая система насекомых с классификацией первичнобескрылых и древнекрылых.
// С.-Петербург: "Лань", 2000. 336 с. WWW
Клюге Н.Ю. 2002. Гомология ротовых частей блох (Aphaniptera). // Энтомологическое обозрение
81 (4): 808–816.
Клюге Н.Ю. 2003. Об эволюции и гомологии генитальных придатков насекомых. // Труды РЭО
74: 3–16.
Darwin Ch. 1871. On the origin of species by means of natural selection, or the preservation of favored races in the struggle for life. 5th edition. New York: D. Appleton and
company. 447 p. BHL
Hennig W. 1950. Grundzuge einer Theorie der phylogenetischen Systematik. // Deutscher Zentralverlag, Berlin. 370 S.
Imms A.D. 1939. On the antennal musculature in insects and other Arthropods. // Quarterly Journal of Microscopical
Science 81: 273–320.
Imms A.D. 1940. On growth processes in the antennae of insects. // Quarterly Journal of Microscopical
Science 81: 585–593.
Kluge N.J. 1999. Mitos en sistematica y principos de nomenclatura zoologica. / Myths in systematics and principles of zoological nomenclature. // Boletin de la Sociedad Entomologica
Aragonesa 26: 347–377.
Kluge N.J. 2004a. The phylogenetic system of Ephemeroptera. // Kluwer Academic
Publishers. 442 p.
Kluge N.J. 2004b. Larval leg structure of Nannochorista and characteristics of Mecoptera. // Russian Entomological
Journal (2003) 12 (4): 349–354.
Kluge N.J. 2005a. Larval/pupal leg transformation and a new diagnosis for the taxon Metabola
Burmeister, 1832 = Oligoneoptera Martynov, 1923. // Russian Entomological
Journal (2004) 13 (4): 189–229.
Kluge N.J. 2005b. Metamorphosis and homology of mouthparts in Neuropteroidea (Hexapoda: Metabola), with remarks on systematics and nomenclature. // Russian Entomological
Journal 14 (2): 87–100.
Kluge N.J. 2007a. Oligoneuria itayana sp.n. (Ephemeroptera, Oligoneuriidae) – a new mayfly species from Peruvian Amazonia. // Russian Entomological
Journal 16 (2): 127–137.
Kluge N.J. 2007b. Review of Ameletidae (Ephemeroptera) of Russia and adjacent lands. // Russian Entomological
Journal 16 (3): 245–258.
Kluge N.J. 2008. A new taxon Hermanellonota, or subtribe Hermanellini subtr.n. (Ephemeroptera, Leptophlebiidae, Hagenulini), with description of three new species from Peruvian Amazonia. // Russian Entomological
Journal (2007) 16 (4): 127–137.
Kluge N.J. 2009a. New version of the database “Ephemeroptera of the World” as the first experience of a permanent and objective web catalogue in biology. // Aquatic
Insects 31, supplement 1: 167–180. WWW
Kluge N.J. 2009b. Higher system of Atalophlebiinae (Leptophlebiidae) with description of three new species of
Terpides s.l. from Peruvian Amazonia. // Russian Entomological Journal
18 (4) 243–256.
Kluge N.J. 2010a. A new autapomorphy of the taxon Tricoryptera and redescription of
Ephemerythus. // Russian Entomological Journal 19 (1): 21–30.
Kluge N.J. 2010b. Redescription of the taxon Tricorygnatha (Ephemeroptera, Tricorythus s.l.) based on new findings in Africa and Indonesia. // Russian Entomological
Journal 19 (2): 79–104.
Kluge N.J. 2010c. Circumscriptional names of higher taxa in Hexapoda. //
Bionomina 1: 15–55.
Kluge N.J. 2010d. Paradoxical molting process in Orthezia urticae and other coccids (Arthroidignatha, Gallinsecta). // Zoosystematica
Rossica 19 (2): 246–271.
Kluge N.J. & Novikova E.A. 2011. Systematics of the mayfly taxon Acentrella (Ephemeroptera, Baetidae), with description of new Asian and African
species. // Russian Entomological
Journal 20 (1): 1–56.
Latreille P.A. 1802. Histoire naturelle, générale et particuliere des Crustacés et des Insectes. T.3. Famille naturelle des
genres. // Paris. BHL
Linnaeus C. 1751. Philosophia botanica. In qua explicantur fundamenta botanica cum definitionibus partium, exemplis terminorum, observationibus rariorum, adjectis figuris aeneis.
// Stockholmiae: apud Godofr. Kiesewetter, 1751. 362 p. BHL
// Русский перевод: К. Линней. Философия ботаники. Москава, "Наука"
1989: 452 с.
Linnaeus C. 1758. Systema Naturae per regna tria Naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. T.I.
// Holmiae: Impensis Direct. Laurentii Salvii: 824 p. BHL
Summary
Cladoendesis is a method of phylogeny analysis opposed to various matrix methods. In contrast to matrix methods, in cladoendesis phylogenetic trees are not build each time as new ones, but are reconstructed on the ground of formerly reached results. Each character of each taxon is compared with its ancestral condition in the ground-plan of the higher taxon. Revealed part of phylogeny is represented in a form of hierarchical classification. In addition to the principles of natural classification, elaborated by C. Linnaeus, P.-A. Latreille and others, evolutionary theory elaborated by Ch. Darwin and others and principles of phylogenetic systematics elaborated by W. Hennig and others, cladoendesis includes rank-free dual nomenclature system (DualNom) and advanced method of text layout elaborated by the author. Important components of cladoendesis are usage of the term "plesiomorpon" and taking into account such important evolutionary events as conservation and deconservation of characters. Cladoendesis allowed the author to make comprehensive revision of phylogeny of Ephemeroptera, find homology in structure of imaginal maxillae of Aphaniptera, larval maxillae of Neuropteroidea-Birostrata, larval legs of Nannomecoptera and genital parts of some insect taxa. It also allows to understand nature and evolution of insect metamorphosis.
It is used to regard that primitively insect ontogenesis passes by the most direct way, while such phenomena as substitutions of organs by means of their atrophy and subsequent restoring, are secondary. In contrast to this, flagellum of antenna of Amyocerata primitively grows by means of adding proximal segments and simultaneous loss of distal segments; in many taxa, including Metabola (insects with complete metamorphosis) this loss of distal segments is lost.
Some authors tried to explain origin of the complete metamorphosis by these or that reasons – ecological, morphogenetic or others. This approach is wrong, because all insects with complete metamorphosis constitute a holophyletic taxon Metabola Burmeister 1832, i.e. originate from a single ancestral species which got this kind of metamorphosis. If the complete metamorphosis could appear by some reason, it would appear, by the same reason, in some other species, and in this case the taxon Metabola, characterized by the complete metamorphosis, would be polyphyletic. Holophyly of Metabola is well proven by cladoendesis, but cannot be revealed by any matrix method. Basing on understanding of these facts, the author was able to discover the concrete features which appeared in the common ancestor of Metabola, and, being conserved, were inherited by all its descendants. Among these features – the loss of scapus in larval antenna (that leads to peculiar transformation of antennae in metamorphosis) and the peculiar kind of leg transformation in course of molt from larva to pupa. During the larval/pupal molt, legs lose musculature and get immobile knee bend, so pupa is unable to use its legs. This conserved feature makes most insects to elaborate inactive mode of life in the pupal stage.
It is used to regard that male coccids (Gallinsecta De Geer 1776) have metamorphosis similar to the complete metamorphosis of Metabola. But phylogenetic position of Gallinsecta and Metabola, as revealed by cladoendesis, does not allow to assume common modifications in their metamorphoses. Now, when concrete autapomorphies of Metabola are known, it becomes possible to compare metamorphoses of Metabola and Gallinsecta. Examination of Orthezia urticae and some other coccids allows to conclude, that metamorphosis of their males has no any modifications common with the complete metamorphosis. Inactive mode of life of their nymphs is connected not with any anatomical reasons, but purely with the fact that in course of transformation from feeding wingless larva to non-feeding winged imago, nymph has already lost mouth apparatus and has not yet got functional wings, so does not need to move. At the same time, actively feeding stages of Gallinsecta, both males and females, have an unusual mode of molt transformation of legs and antennae; this is a unique autapomorphy of Gallinsecta, not found in any other insects.