 I.4. Reconstructing phylogeny
I.4. Reconstructing phylogeny| nomenclature | general system and phylogeny of insects | systematics of Ephemeroptera |
| last update 16.V.2022 |
PRINCIPLES OF CLASSIFICATION OF SUPRASPECIES TAXA
|
The division I.1. from the book by N. Kluge |
I.4. Reconstructing phylogeny
 I.5. Principles of classification of supraspecies taxa
I.5. Principles of classification of supraspecies taxa
I.5.1 Mono-, poly-, para-, and holophyly
I.5.1.1 Background
I.5.1.2 Definitions
I.5.1.3 Holo-, para- and polyphyly: explanation of terms
I.5.2 Various approaches to classification
I.5.2.1 Cladism, of phylogenetic systematics
I.5.2.2 Traditionalism, or evolutionary systematics
I.5.2.3 Gradism
I.5.3 Classification principles in terms of cladoenedesis
I.5.3.1 Benefit and damage from paraphyletic (ancestral) taxon
I.5.3.2 Plesiomorphons
I.5.3.3 Aspiration of classification for the cladistic ideal
I.5.3.4 Reasons of disagreements between cladists and traditionalists
I.5.4 Relative and absolute ranks of taxa
I.5.4.1 Strict hierarchy of classification
I.5.4.2 Absolute ranks
I.5.4.3 Inequality of absolute ranks
I.5.4.4 Problem of genus
I.6. Principles of nomenclature of zoological taxa
I.5.
Overwhelming majority of investigators agree that classification should reflect phylogeny, as only phylogenetic connections between animal species can serve as a base for their natural classification. However, accepting this thesis, it is necessary to understand clearly that the classification can not be identical with the phylogeny. This statement does not mean that the classification obligatory should reflect something other besides the phylogeny (while some authors do have such opinion – see I.5.2.2 and I.5.2.3). We can build a classification which reflects phylogenetic connections only (as it is postulated by cladists – see I.5.2.1), but in any case the classification is not identical with phylogeny, in the same manner as any reflection is not identical with the object which it reflects. In the classification we use artificial signs to reflect that aspects of the phylogeny which we are able to discover; the phylogeny itself exists independently from our mind and independently of our ability to discover it.
Different authors suggest and state different principles of uniting animals to taxa. At the present time, the most known are cladistic and traditionalistic principles, they are discussed below. Besides them, we can name a phenetic, or numerical principle of classification, which was used for a short time by narrow circle of authors and did not justified itself (see I.4.2.6.3.B), and a gradistic principle, which is not popular among recent systematicists (see I.5.2.3). While there are various opinions how to establish taxa, the terms which characterize taxa in relation to a phylogenetic tree are understood universally – these are first of all the terms holophyly, paraphyly and polyphyly.
I.5.1. MONO-, POLY-, PARA-, AND HOLOPHYLY
Depending on phylogenetic nature of taxa, the taxa are divided to monophyletic and polyphyletic, or to holophyletic, paraphyletic and polyphyletic.
I.5.1.1. Background
The terms «monophyly» and «polyphyly» were introduced by E. Haeckel in XIX century and are widely used since that time, but during long period they had no distinct definitions; as a result of this different authors suggested different definitions of these terms, which sometime contradict one another. There was widely accepted a definition of monophyletic taxon as «arising from one common ancestor», and of a polyphyletic taxon as «arising from several different ancestors». However, if proceed from the fact that all organisms arose from one common ancestor, the definition of monophyly as «arising from one common ancestor» should be regarded to be not enough, because in this case every taxon would appear to be «monophyletic». W. Hennig gave his own strict definition of the term monophyly (see below the definition of holophyly, or monophyly according to Hennig), but at the same time he restricted meaning of the term «monophyly» in comparison with that meaning which is intuitively understood by majority of authors. Because of this, other alternative definitions were suggested.
Some authors (Simpson 1961, Mayr 1969) defined monophyletic taxon as a taxon arising as one or more lines from one directly ancestral taxon of the same or lower rank. It is clear that such definition is absolutely useless, because the main concept here is a rank of the taxon which is an artificial category (see I.5.4 below) and, hence, the concept of monophyly becomes to be artificial. If accept such definition, any set of species can be regarded as a «monophyletic» taxon, if attribute a corresponding rank to it.
For example, if insects with various independently arisen sucking mouth apparatus would be united in a single taxon Haustellata Clairville 1789 – in this case according to the generally accepted idea about mono- and polyphyly, the taxon Haustellata would be surely polyphyletic; however, if at the same time we would unite all other insects in a taxon Mandibulata Clairville 1798, and would attribute equal ranks to the taxa Haustellata and Mandibulata (for example, would regard them to be subclasses, as it was done in some papers of the end of XVIII – beginning of XIX century) (see Chapter VI: Pterygota: «Classifications» I) – in this case according to the Simpson's definition, the taxon Haustellata turns from polyphyletic to a «monophyletic» one, while its composition and our opinion about its origin are not changed.
Available definitions of monophyly and polyphyly, which from one side, are distinctly formulated, and from other side correspond to traditional understanding of these terms, were suggested by Ashlock (1971) (see below). Together with these definitions, in order do distinguish the wide monophyly and the monophyly according to Hennig, Ashlock suggested to use a new term «holophyly» instead of «monophyly» according to Hennig. The term «paraphyly» was introduces by Hennig and is always used only in its original meaning (see below).
I.5.1.2. Definitions
All taxa are divided into monophyletic (in wide sense) and polyphyletic ones.
1. Monophyletic taxon (monophylum). Monophyletic in wide sense is a taxon which includes an ancestor common for all members of this taxon, and also includes all phylogenetic branches going from this ancestor to each of the members of this taxon. Under the words «a common ancestor» we understand at least the nearest one among that ancestral species from which all other species of the given taxon had originated. Usually the common ancestor of the taxon, as well as other ancestral forms, is absent in recent fauna and is unknown to investigators; we regard that the ancestor agrees is included in the given taxon if its assumed characters agree with the diagnosis of this taxon.
Monophyletic (in wide sense) taxa are divided into holophyletic (or monophyletic in narrow sense) and paraphyletic ones.
1.1. Holophyletic taxon (holophylum) is the same as strictly monophyletic, i.e. monophyletic according to Hennig. Holopyhyletic is a taxon which includes not only its common ancestor and all phylogenetic branches going from this ancestor to each member of this taxon, but also includes all descendants of this ancestor. In the Fig. 1.2A taxa a-b-c-d-e and b-c-d are holophyletic ones. Among characters which characterize a holophyletic taxon, there are autapomorphies of this taxon, which at the same time are synapomorphies of all species included into this taxon. Indeed, an apomorphy which have originated in the ancestor of a holophyletic taxon, can be retained by all its descendants (i.e. in the whole taxon), and since no any descendants of this ancestor occur outside of this taxon, this apomorphy exists in this taxon only. From the definition proceeds, that a holophyletic taxon can not be ancestral to any other taxon, because all forms originated from members of the holophyletic taxon, are included into this taxon. If we would outline a holophyletic taxon by a closed line on a phylogenetic scheme, this line will cross the phylogenetic tree only in one place – at the entrance to the taxon. In the term holophyly the first base «holo-» (from Greek ολο-, integral) means that the taxon is formed by an integral branch of the phylogenetic tree; this branch is cut from the remain tree in a single place (which represents the base of this branch) and is included in this taxon together with all its branchings.
1.2. Paraphyletic taxon (paraphylum). Paraphyletic is a taxon which includes an ancestor common for all members of this taxon, and all phylogenetic branches going from this ancestor to each member of this taxon, but at the same time includes not all descendants of this ancestor. The paraphyly is a form of the monophyly in wide sense, but it differs from the monophyly after Hennig, i.e. from the holophyly. In contrast to a holophyletic one, a paraphyletic taxon is ancestral to some other taxon (or several other taxa) and differs from it (or from them) by plesiomorphies only. In the Fig. 1.2B, a paraphyletic taxon a-b-d-e is shown, which is ancestral for the taxon c. If we would outline a paraphyletic taxon by a closed line on a phylogenetic scheme, this line will cross the phylogenetic tree more than in one place – at the entrance to the taxon and at the exit (or exits) from it.
2. Polyphyletic taxon (polyphylum). Polyphyletic is a taxon which does not include any ancestor common for all members of this taxon, or does not include some portions of phylogenetic branches uniting the common ancestor of the taxon with some of its members (it would be more exactly to say that these ancestors do not correspond to the diagnosis of the taxon). Characters on which diagnosis of a polyphyletic taxon is based, originated in evolution several times independently. In the Fig. 1.2C polyphyletic taxon a+b is shown. If we would outline a polyphyletic taxon by a closed line on a phylogenetic scheme, this line will cross the phylogenetic tree more than in one place – not less than two times at different entrances to the taxon.
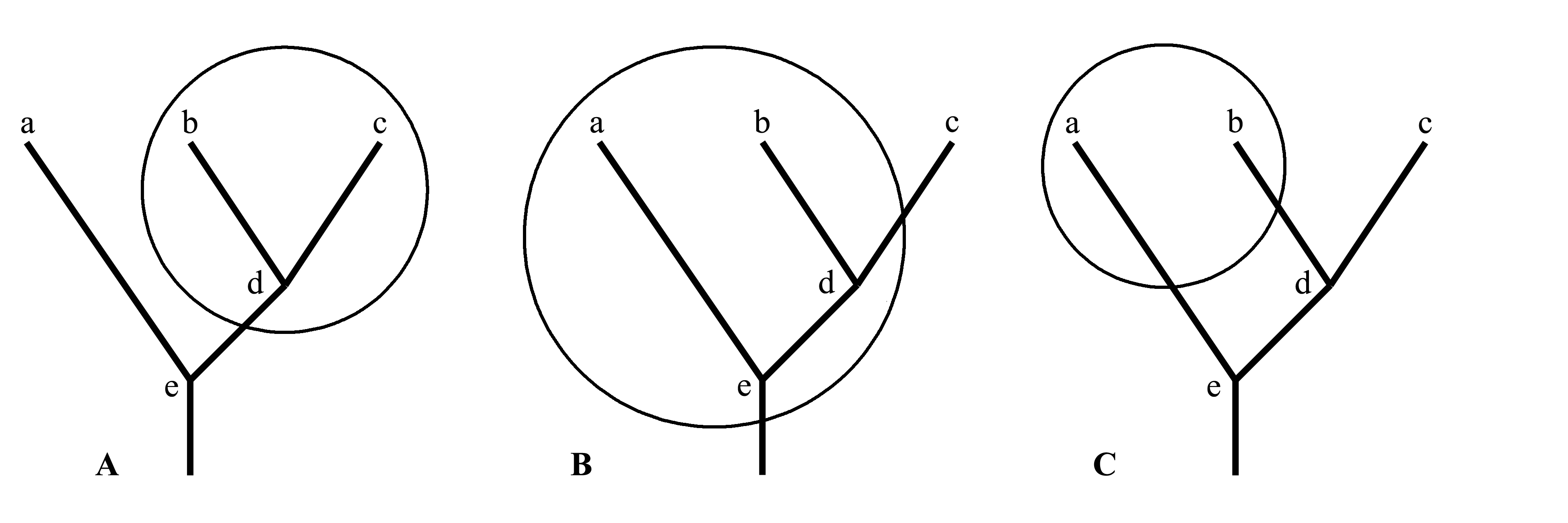
Fig. 1.2. Three ways to establish a large taxon in one and the same group of animals consisted of three recent taxa a, b and c and ancestral forms d and e. Phylogenetic relations are shown by thick line, the taxon inder consideration is outlined by a ring: A – holophyletic taxon (b+c+d); B – paraphyletic taxon (a+b+d+e); C – polyphyletic taxon (a+b).
In order to avoid comments «in wide sense» and «in narrow sense» it is possible not to use the term monophyly which has two meanings, but to use only the terms «holophyly», «paraphyly» and «polyphyly», each of which is has a single meaning.
I.5.1.3. Explanation of the terms holo-, para- and polyphyly
In a shortest manner, the definitions of these categories can be expressed by the following scheme:
|
The taxon under consideration: |
Includes
all |
Includes
not all |
|
Includes its common ancestor |
holophyletic |
paraphyletic |
|
Does not include its common ancestor: |
polyphyletic |
|
Sometimes wrong understanding of these terms leads to confusion. Here we will pay attention to that points which sometimes produce difficulties in understanding of the terms holophyly, paraphyly and polyphyly.
Firstly, it is necessary to remember that these terms are used only applying to taxa, but not to organisms and not to phylogenetic branches. Because of this, before starting discussion if the given taxon is holo-, para- or polyphyletic, it is necessary to outline distinctly this taxon, i.e. to decide which species are included into it and which not. Usually in order to establish boundaries of the taxon, a diagnosis is used: in the diagnosis a certain set of characters is listed, and it is assumed that if characters of an animal correspond to this diagnosis, this animal is included into this taxon, and if do no correspond – is not included. If boundaries of the taxon are not established, the concepts «include» – «does not include» (ancestor or all descendants) lose their meaning and the definitions become unavailable.
Establishing of taxon boundaries is an artificial procedure, as well as the concept of taxon itself. When we have artificially established a taxon with its boundaries, we can ask a question, if this taxon is natural, i.e. in how does it correspond to a natural concept – phylogenetic branch, or by other words, is it a holophyletic, paraphyletic or polyphyletic taxon. If change boundaries of the taxon, the answer on the question if this taxon is holo-, para- or polyphyletic, will be also changed. Here we regard that if we have changed boundaries of the taxon, we have finished to discuss the previous taxon and starts to discuss a new taxon. However if use a rank-based nomenclature, a taxon is regarded to retain itself when its boundaries are changed, i.e. the same name is attributed to taxa with different boundaries (see I.6 below), that sometimes leads to confusion.
For example, the genus Phryganea Linnaeus 1758 originally included various amphibiotic insects (caddisflies and stoneflies) and was a polyphyletic one; recently taxon of such circumscription is not recognized by anybody; but in correspondence to the rules of zoological nomenclature, the generic name Phryganea L. is retained and recently is applied to a taxon of much smaller circumscription, which is holophyletic.
Secondly, an answer to the question if the taxon is holo-, para-, or polyphyletic, can be given only in connection with a certain hypothesis about phylogeny of this group of organisms.
For example, if we accept the hypothesis about close relationship of Onychophora, Myriapoda and Hexapoda (see scheme in I.4.1.5), in this case the taxa Gnathopoda, Euarthropoda and Mandibulata would be regarded by us as polyphyletic, and the taxon Ceratophora as a holophyletic one; but if we would accept the hypothesis about close relationship of Eucrustacea, Myriapoda and Hexapoda (see ibid.), we would think that the taxa Gnathopoda, Euarthropoda and Mandibulata are holophyletic, and the taxon Ceratophora – a polyphyletic one. Actually for each taxon only one answer is correct, and the aim of each biologist is to find it reconstructing phylogeny.
Thirdly, the concepts holophyletic, paraphyletic and polyphyletic taxon are absolute concepts, in contrast to the concepts apomorphic and plesiomorphic characters, which are relative ones. Indeed, a character can be apomorphic only relatively to another alternative character, which is plesiomorphic relatively to the given character: a character B can be apomorphic relatively to a character A, but plesiomorphic relatively to character C (see I.4.2.1). As for a taxon, it is holophyletic, paraphyletic or polyphyletic not relatively to this or that other taxon, but generally speaking. In the definitions of holo-, para-, and polyphyly, only natural concepts «ancestor» and «descendants» are reported, but not other taxa apart of the taxon to which the definition is applied. For example, if any descendants of a member of a taxon N are not included into the taxon N, the taxon N is termed paraphyletic independently in which manner we group into taxa that descendants of N which are excluded from N.
Fourthly, the concepts holo-, para-, and polyphyletic taxon are independent not only from classifications of other taxa, but from characters as well. Since the circumscription (i.e. boundaries) of a taxon are established usually with help of characters (which form a diagnosis), and the phylogeny is reconstructed usually on the base of apomorphies (which are characters again), a wrong impression appears that all these concepts are based on characters. However, as wrote Linnaeus, «not a character determines a genus, but a genus determines a character» – by other words the characters only help us to discover taxa and phylogeny, but do not determine them. Phylogenetic relationships exist in nature independently if we can reconstruct them with help of cladistic analysis (which is an analysis of characters) or not. In the same manner, if a taxon includes its common ancestor and all its descendants, this taxon is holophyletic independently if its diagnosis contains autapomorphies or not; if its diagnosis contains autapomorphies, they serve for us as a proof of its holophyly; if autapomorphies are absent or unknown, we do not know if this taxon is holophyletic or not, but such our non-acquaintance does not make the taxon to stop being holophyletic. The presence of apomorphies in a taxon diagnosis is not a definition of the holophyly, but only a way to clarify the holophyly; but this way of clarifying holophyly is the only known one, and because of this somebody erroneously thinks that this is a definition.
Fifthly, it is necessary to specify what concretely is understood under the term «ancestor» in the definitions of monophyly and polyphyly. Each organism has a lot of ancestors, chain of which ascends to it from the origin of the life on the Earth. In its turn, each taxon has a lot of common ancestors, among which is the whole chain of ancestors from the origin of the life up to the first divergence which leaded to separation of any members of this taxon. Thus no one taxon can include inside itself all its ancestors. The definition of the monophyly (as well as the definitions of the holophyly and the paraphyly) states that the taxon includes any ancestor, at least the nearest one among the ancestors common for all members of this taxon. Correspondingly, a polyphyletic taxon includes no one ancestor which would be common for all members of the given taxon.
I.5.2. Various approaches to classification
I.5.2.1. Cladism, of phylogenetic systematics
Here and below, the term «cladism» is applied to scientifically grounded principles of classification, but not to formal numerical and matrix methods widely distributed in modern biological literature (about their discrepancy to scientific principles – see I.4.2.6).
Cladism is the only consistent and distinctly formulated principle of classification. However, its positions look paradoxical, because of which some authors do not recognize it. As shown below (see I.5.3), it is impossible to built a classification which would completely correspond to demands of the cladism, but the whole development of the systematics is an approach to such classification.
Principles of cladism were formulated by Willi Hennig (1950), and the author termed these principles phylogenetic systematics. The word composition «phylogenetic systematics» is not quite successful, because similar word compositions «systematic phylogeny» and «phylogenetic classification» were used in the classical papers by Haeckel (1896, 1898). The term cladism is formed from the Greek word κλαδος – branch. In the cladistic systematics taxa represent themselves branches of the phylogenetic tree; it can be also said that the cladistic classification is a classification of phylogenetic branches, but not of any other natural objects.
The main essence of the cladism can be formulated very shortly: all taxa should be only holophyletic (i.e. monophyletic according to Hennig). Neither polyphyletic, nor paraphyletic taxa are allowed in the classification.
According to this principle, the hierarchy of taxa turns out to be uniquely corresponding to the branching hierarchy of the phylogenetic tree. Thanks to this, arbitrariness in the construction of the classification is eliminated. Indeed, if all taxa should be only holophyletic, there is no choice between different ways of dividing the phylogenetic tree into taxa: in all cases a boundary between taxa of the highest rank should pass by the earliest divergence of the phylogenetic tree. The principle of cladism leads to important consequences.
1) If all taxa are holophyletic, taxa never can be maternal (ancestral) and daughter, they can be only sister or non-sister.
Indeed, every taxon ancestral to another (non-subordinated to it) taxon, is paraphyletic according to the definition, and hence, should not exist. Here we must not mix concepts of ancestor and ancestral taxon: existence of ancestors is recognized by cladists in the same manner as it is recognized by all other evolutionary thinking biologists (only people with very extravagant opinion think that living organisms can originate not from their parents, but in some other manner). In contrast to the ancestor, the ancestral taxon can be created by us or not; apologists of the cladistic systematics regard that the ancestral taxon should not be created.
2) Not all species can be attributed to taxa of all ranks. Indeed, in Fig. 1.3 the species 3, which is a common ancestor of Lepidoptera (moths and butterflies) and Trichoptera (caddisflies) can not be attributed neither to caddisflies (because in this case the taxon Trichoptera would become paraphyletic), nor to butterflies (because in this case the taxon Lepidoptera would become paraphyletic). For this species 3 a separate taxon (opposing both to Trichoptera and Lepidoptera) also can not be established, because in this case it would be a paraphyletic taxon. Thus, the common ancestor of caddisflies and butterflies does not belong neither to Trichoptera, nor to Lepidoptera, nor to another taxon of the same rank, but together with caddisflies and butterflies belongs to a higher taxon Amphiesmenoptera.
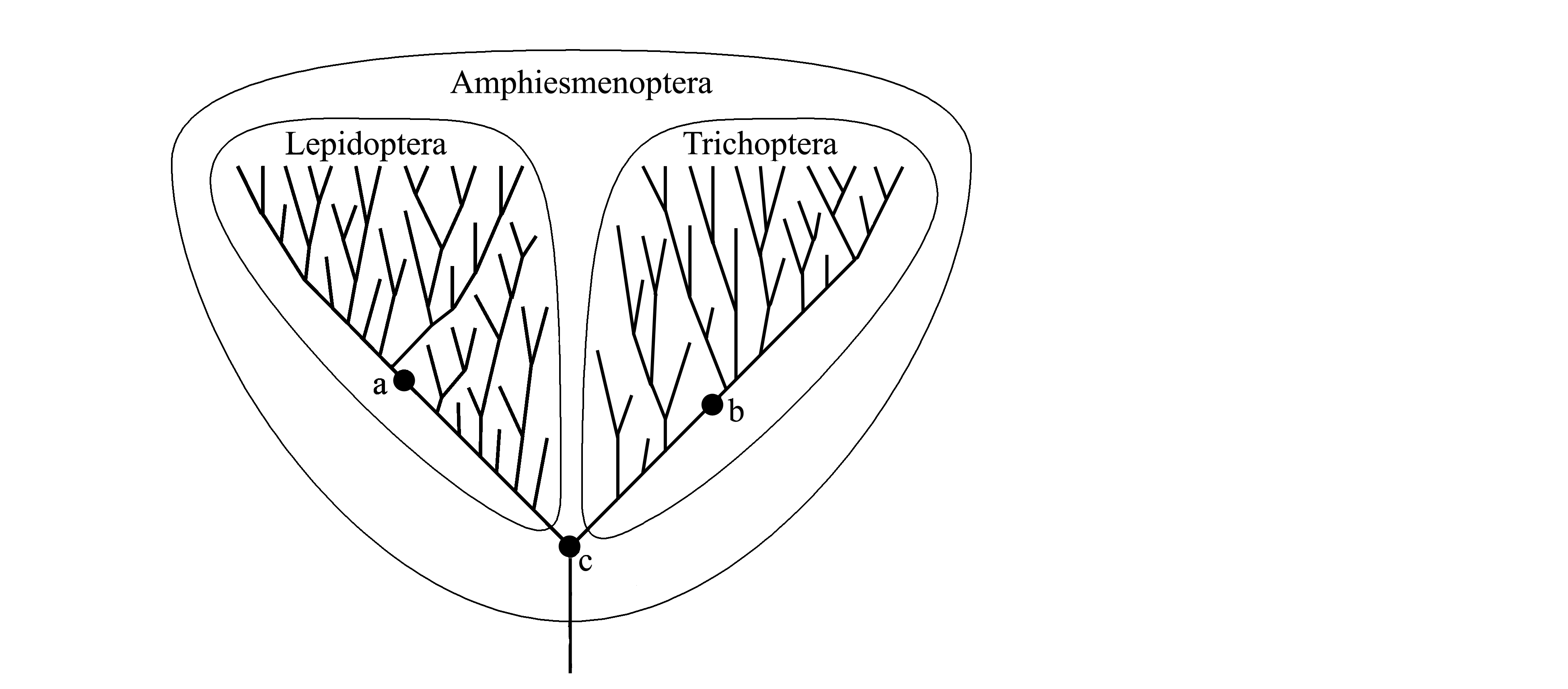
Fig. 1.3. Boundaries of the taxa Lepidoptera, Trichoptera and Amphiesmenoptera in cladistic systematics. Phylogenetic branches are arbitrarily shown by thick line; boundaries of taxca are outlined by thin line; thick dots indicate the following ancestral species: a – common ancestor of all recent Lepidoptera; b – common ancestor of all recent Trichoptera; c – common ancestor of all Lepidoptera and Trichoptera.
If not to follow the cladistic principle, but to establish paraphyletic taxa in order to include all species into taxa of all ranks, it appears to be unclear where to make boundaries between the taxa. Indeed, as on a phylogenetic tree natural boundaries of taxa are absent (because the whole tree represents a non-interrupted branching chain of generations – Fig. 1.1), in each concrete case it is difficult to come to agreement where an artificial boundary between taxa should pass. In the cladism a universal answer on this question is suggested: the boundary between two sister taxa should pass by their common ancestral species, and the ancestral species itself does not belong to anyone of these taxa.
About real usage of the cladistic principles see I.5.3 below.
I.5.2.2. Traditionalism, or evolutionary systematics
The traditionalism appeared as an opposition to the cladism, and its aim is to protect and ground that positions which traditionally exist in systematics, but are rejected by the cladism. The ideas of traditionalism were formulated by Mair (1971) and others. In the recently existent classifications of living organisms, not only holophyletic, but wittingly paraphyletic taxa are present as well. According to opinion by traditionalists, the paraphyletic taxa not only can, but must exist along with holophyletic taxa. As for polyphyletic taxa, they should be absent; in this point opinions of traditionalists and cladists coincide. It is stated that a natural taxon should be first of all characterized by characters inherited from the ancestor; such characters are not only apomorphies (which characterize a holophyletic taxon), but plesiomorphies as well (which characterize a paraphyletic taxon). In connection with this, in traditionalism a concept of ancestral taxon is present – i.e. related taxa of the same rank can be not only sister ones, but maternal-daughter as well.
While cladists regard that in order to make a unique classification it should be built on the base of a single parameter – the succession of divergences of the phylogenetic tree (i.e. cladogenesis), according to the opinion of traditionalists the classification should be built on base of two parameters – 1) cladogenesis, i.e. succession of divergences of the phylogenetic tree, and 2) anagenesis, i.e. a degree of evolutionary changes in each branch (here the term anagenesis is used in wide sense, as it was defined by J. Huxley 1957). Graphically this difference is usually illustrated as in Fig. 1.4.
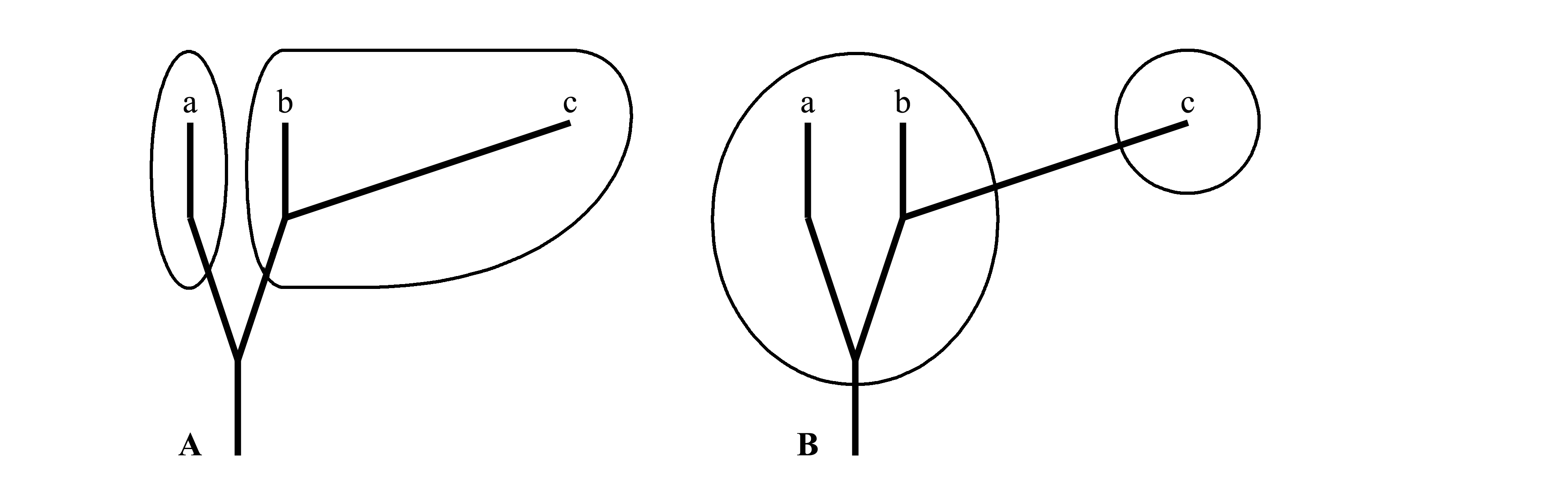
Fig. 1.4. Two variants of division of one and the same group of animals, consisting of three recent taxa a, b and c, to two large taxa. Phylogenetic branches are shown by thick line; taxa bounaries are outlined by thin line; highth of branches reflects geological time; horisontal length of branched shoul reflect amount of evolutionary changes. A – cladistic classification: each of two taxa (the taxon a and the taxon b+c) is holophyletic. B – traditionalistic (evolutionary) classification, the taxon a+b is paraphyletic, the taxon c is holophyletic.
In the Fig. 1.4A, the group is divided to two taxa according to the cladistic principle, i.e. by the oldest divergence and in the Fig. 1.4B, the same group is divided in different manner, in correspondence with the traditional principle, which takes in account anagenetic component which is bigger in the branch c.
At the same time it is unclear how is it possible to determine objectively a quantity of evolutionary changes (i.e. the length of branches on drawing). Traditionalists do not recognize poorly numerical evaluation of degree of difference (which is accepted in phenetics); in contrast to pheneticists, they attribute to different characters different weight and take into account not only degree of difference, but also a succession of divergences of the phylogenetic tree.
Since in the traditionalism two parameters are used at the same time, it is unclear which of them in which cases should be preferred. Because of this, in limits of the traditionalism different classifications can be done on the base of the same phylogenetic tree: for example, in the case in the Fig. 1.4, one can regard that the difference between c and a+b is more important than the succession of divergences, and on this base chose the classification B; or on the contrary, regard this difference less important and on this base chose the classification A.
About real usage of the traditionalistic principles see I.5.3 below.
I.5.2.3. Gradism
In contrast to the cladistic and traditionalistic schools in systematics, each of which pretends to be the only one and to serve as the basis for the whole systematics, gradism usually is not regarded as a universal principle of building classification. A place of gradism among other approaches to systematics is that some authors regard to be appropriate to have gradistic taxa along with taxa established according to other principles.
It can be figuratively said, that in contrast to the cladistic classification where the phylogenetic tree is cut to taxa along, in the gradistic classification it is cut to taxa across. Such taxa, termed also grades, represent levels of organization and can be either holophyletic, or paraphyletic, or polyphyletic (Fig. 1.5).
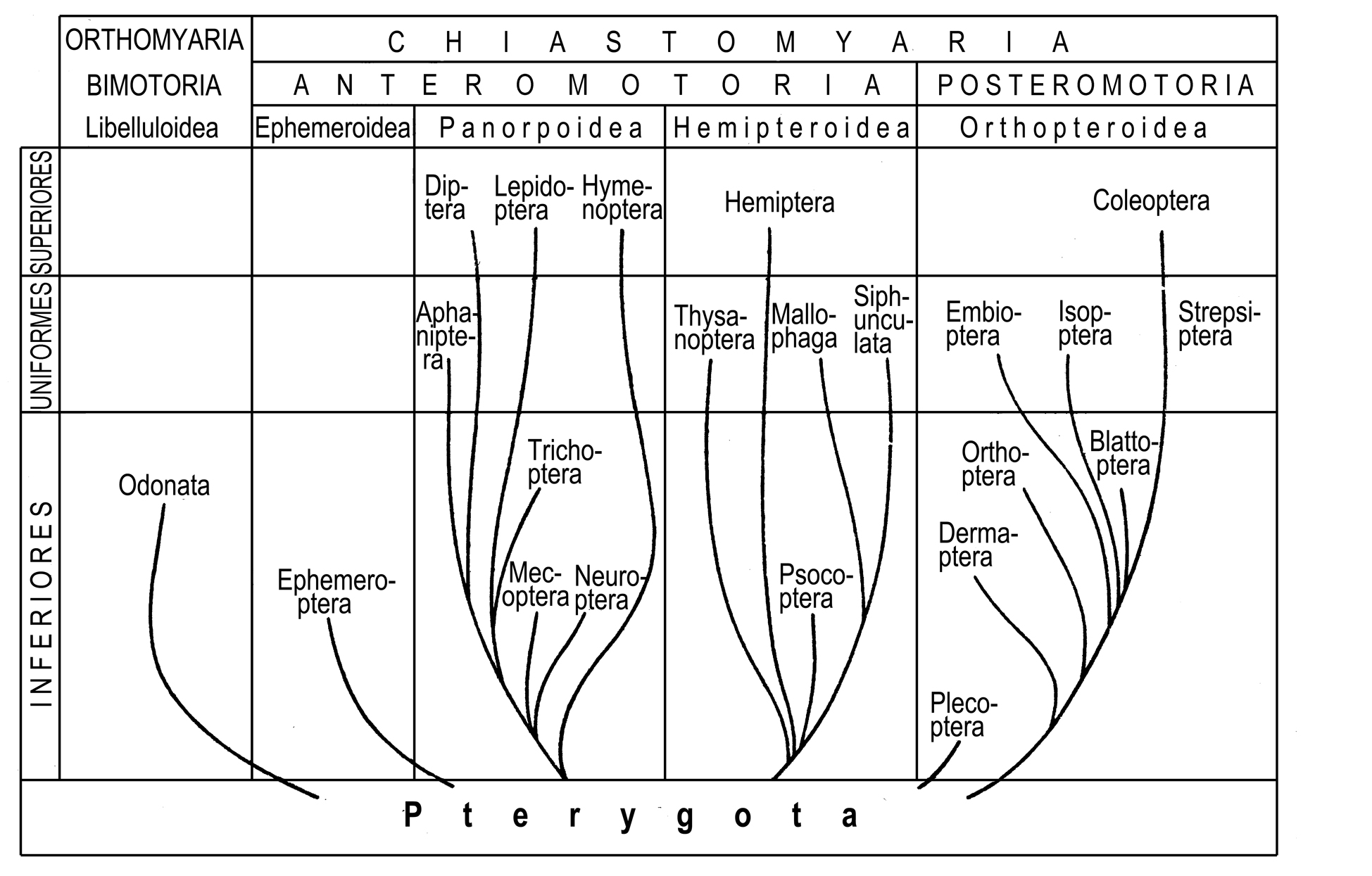
Fig. 1.5. Gradistic classification of winged insects by Schwanwitsch (1946).
With this, here is no distinct definition what should be regarded to be different levels of organization. One can arbitrarily choose any of the characters acquired in evolution, calling them transitions to new levels of organization; depending on this choice, different classifications will be obtained based on the same phylogenetic tree.
Presence of polyphyletic taxa in gradistic classifications is determined by existence of convergence: diagnoses o these taxa are based on independently appeared characters. If there were some regularity in convergences, and a presence of convergence in certain character would allow to make some prognosis, then a classification based on convergences would make sense. However, at the present time such regularities are unknown and it is unclear if they exist at all. Therefore, the gradistic principle of classification is not widely accepted, in contrast to the cladistic one (see above), where taxa being corresponding to phylogenetic branches, are theoretically grounded and have evident prognostic value.
I.5.3. principles of Classification in terms of cladoenedesis
As it was shown above (I.5.2), different authors declare different principles of building classification. However, that principles which are declared, not always correspond to principles which the same author actually uses when builds a classification.
The expediency of existence of holophyletic taxa and inexpediency of polyphyletic taxa is recognized both by cladists and traditionalists. Disagreements are connected with a question about expediency of paraphyletic taxa: cladists do not recognize them, and traditionalists do recognize.
Before discussion of the principles of cladoendesis, let us look what significance have paraphyletic taxa.
I.5.3.1. Benefit and damage from paraphyletic (ancestral) taxon
If to be exact, an ancestor of any taxon of bisexual organisms can be only species or a smaller set of specimens (subspecies, race, population) inside which free redistribution of genetic information takes place; in organisms which have no sexual reproduction, this is a selected specimen. However a species ancestral for the taxon usually is absent in recent nature, in fossil conditions unknown, and we have no enough information to reconstruct it as exact as species characters. We can reconstruct the ancestor interesting for us as exact as characters of a taxon of higher rank (genus, family, order, class, phylum and so on).
If existence of paraphyletic supra-species taxa is allowed, instead of ancestral species we can speak about an ancestral or maternal (and hence paraphyletic) genus, family, order, class, phylum and so on). In this case it is necessary to remember that actually the ancestor of the taxon is not the whole ancestral supra-species taxon, but only one species of this taxon which is unknown to us but which really existed in the past. Usage of the term «ancestral taxon» sometimes leads to confusions. Some people think that if a certain supra-species taxon is called «ancestral», this means that different species of this taxon evolved in the same direction (presumably coming to an agreement one with another in some inexplicable manner) and in such way formed a sister taxon of supra-species rank.
For example, if the class Hexapoda is divided to subclasses Apterygota (primarily wingless insects) and Pterygota (winged insects) (see Chapter VI: Hexapoda: «Classifications» I), we can say about the subclass Apterygota, that it is paraphyletic and ancestral for Pterygota, or what is the same, Apterygota is more primitive than Pterygota. On base of this statement, some people seriously thought that different representatives of Apterygota got wings and turned to these or that representatives of Pterygota. Beside this, all species attributed to the ancestral, i.e. the primitive taxon, get a stamp of «primitive» ones. Because of this curious confusions appear; for example an entognathous mouth apparatus of Entognatha, which actually represents a deep modification, was regarded by some authors as a «primitive» condition ancestral to ectognathous mouth apparatus of other insects, only on the base of the fact that insects which have the entognathous mouth apparatus, are united with a primitive ancestor of winged insects into the primitive taxon Apterygota.
The statements «Apterygota gave origin to Pterygota» or «Pterygota originated from Apterygota» appear to be wrongly understood because here the words Apterygota and Pterygota having the same form (the both are names of subclasses, both in plural) must be differently interpreted. Under the word «Apterygota» here should be understood «some of representatives of the taxon Apterygota», but never «any representative of Apterygota»; at the same time, the word «Pterygota» means «any representative of Pterygota».
This difference may be clear to someone who establishes a paraphyletic taxon or expresses such statement, but is is not always clear to someone who accepts these statements.
Thus, for example, usage of a paraphyletic concept «monkey» in the sentence «a man originated from a monkey» have made this sentences a famous target for attacks and jeer by opponents of the evolutionary theory and opponents of science in general. Some people which regard themselves to be evolutionists, finding in this sentence words of the same form «man» and «monkey» believe that if here the word «man» means «every man», thus the word «monkey» should mean «every monkey». As a result of this confusion, the scientific theory about origin of man transforms to an absurd version, that if give a stick in hands of any monkey, and to create for it some other mysterious conditions, by force of some inexplicable «laws of evolution» after many generations descendants of this monkey will become indistinguishable from people. Of cause, hearing such version, somebody who is not highly educated but is able for critical thinking, feels disappointment about science.
To the question «Does modern science claim that man originated from monkey?» it should be answered that modern science cannot claim this, because it does not have a term corresponding to the word «monkey». Indeed, the classification by C. Linnaeus (1758) had both the genus Homo (man) and the genus Simia (monkey); the Latin names of both these taxa are formed in a single form and, being the names of the generic group, are used in the singular (see I.6.3.1.4), so they are translated exactly as «man» and «monkey». Using this classification, we could adequately express modern evolutionary ideas by the words «man (i.e. the holophyletic genus Homo) originated from a monkey (i.e. from the paraphyletic genus Simia)». However, in the modern classification of animals, there is neither the genus Simia in the understanding of Linnaeus, nor a taxon of any other rank corresponding to it in composition. For the species belonging to the Linnaean genera Simia and Homo, Haeckel (1866) established the order Simiae, divided into the suborders Arctopitheci, Platyrrhinae and Catarrhinae (since these taxa are not genera, they are assigned non-typified names in the form of noun plurals; in contrast to generic names, they are not written in italics). Within Catarrhinae, it is customary to distinguish the superfamilies Cercopithecoidea and Hominoidea; the latter is divided into the families Holobatidae and Hominidae; the latter is divided into subfamilies Ponginae and Homininae; the latter is divided into the genera Gorilla, Pan and Homo. Here, the genus Simia in the understanding of Linnaeus corresponds to the totality of all species of the taxon Simiae except for the genus Homo. In this classification, each taxon is holophyletic and cannot be ancestral for any other taxon, including the genus Homo: the taxa Simiae, Catarrhinae, Hominoidea, Hominidae and Homininae include the genus Homo as a part, and the taxa Platyrrhinae, Cercopithecoidea, Holobatidae, Ponginae, Gorilla and Pan are related, but not ancestral to man.
Hence, the question if paraphyletic taxa are useful or not, does not have an unambigous: existence of this taxa allows us to formulate easily ideas about relations ancestor-descendant, but easily formulated idea is not always easily and adequately understood by others.
I.5.3.2. Plesiomorphons
Really existent classifications, as usual, lack wittingly paraphyletic taxa (as well, as they lack wittingly polyphyletic ones), but include only plesiomorphons and wittingly holophyletic taxa. Plesiomorphon is a taxon which is characterized by plesiomorphies only. Since chararacteristic of plesiomorphon does not contain autapomorphies, its holophyly is not proven and not assumed. At the same time, it would be not grounded if call such taxon paraphyletic, because its paraphyly is also not proven. In order to prove holophyly of a taxon a+b on the Fig. 1.7A, it is necessary to discover a synapomorphy common for one of its parts (b) and another taxon (c). If such synapomorphy will be discovered, the taxon a+b could be justifiably called paraphyletic; but at the same time there will appear a possibility to break it, uniting one of its parts b with the taxon c into a new holophyletic taxon b+c, which will be characterized by the newly discovered apomorphy. Thus, a plesiomorphon can be in fact a paraphyletic taxon, but it exists in classification only until its paraphyly is not proven. Some of plesiomorphons can appear to be holophyletic taxa.
The term «plesiomorphon» (in plural «plesiomorphons») was recently introduced (Kluge 2004a) instead of the wrongly used term «plesion». Initially the term «plesion» was introduced not in order to indicate status of a taxon, but in order to indicate place of a taxon in a rank-free hierarchical classification; such plesion can be either wittingly holophyletic, or paraphyletic (Patterson & Rosen 1977). In the previous edition (Kluge 2000) the term «plesion» was wrongly used to indicate a taxon which is not wittingly holophyletic – i.e. for a plesiomorphon. Some authors, in order to indicate plesiomorphon, put a corresponding taxon name into commas (Patterson & Rosen 1977). This is designation is unfortunate, because the same commas are used to indicate wrong or doubtful taxon name, independently of status of the taxon itself.
I.5.3.3. Aspiration of classification for the cladistic ideal
As knowledge is accumulated, the classification permanently changes aspirating to such an ideal, which is the cladistic classification; the actual classification always approaches this ideal, but never reaches it.
A following rule of classification change can be formulated : the classification always changes in such a manner that ratio of the total volume of all holophyletic taxa increases in relation to the total volume of all paraphyletic taxa. This process is well traced if take as an example change of classification of any group of animals during any period of time, both in pre-evolutionary period and evolutionary period of systematics development.
Some authors regard that there is a deep difference between the cladistic and the traditionalistic systematics, because the cladism allows existence of holophyletic taxa only, and the traditionalism – both holophyletic and paraphyletic ones. Actually difference here can be only in the principles declared, but not in the results of building of classification.
Each cladistic taxon (which according to the definition should be holophyletic, i.e. should represent a phylogenetic branch) actually can represent not any branch, but only such phylogenetic branch which is known to us; we can know only such branch which could be reconstructed with help of the phylogenetic analysis (because other methods of phylogeny reconstruction are unknown); the phylogenetic analysis is possible only if the branch has an autapomorphy, which is a concrete character. Thus in the cladistic systematics each taxon is obligatory characterized by a character, while in nature not each branch has its own character. Some phylogenetic branches can have no apomorphies at all, and in this case at the present level of our knowledge we can not reveal this branch and, hence, can not attribute to this branch a taxon status. If in the Fig. 1.6 no any evolutionary changes took place in the segment between E and D, the branch A+B has no any apomorphies, and because of this its existence can not be proved, and hence, a taxon can not be established for this branch.
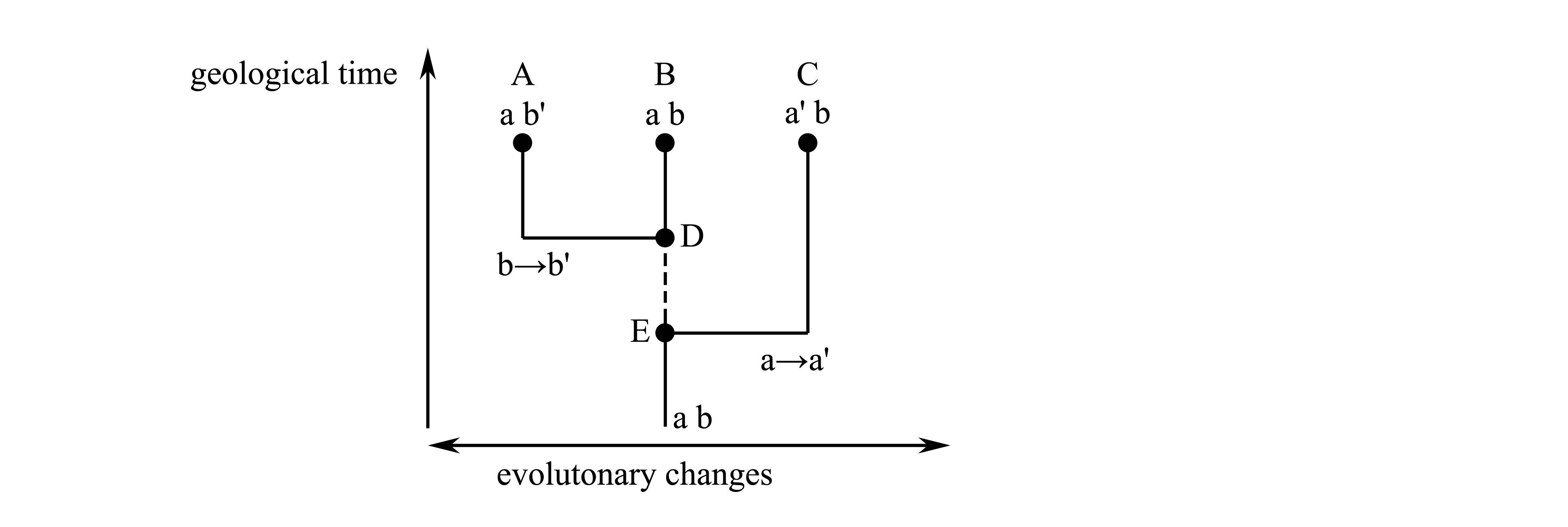
Fig. 1.6. Phylogenetic relationships of taxa, which exist in nature, but can not be reconstructed. A, B and C – recent taxa; D and E – ancestral forms; a and a', b and b' – characters, among which a' and b' – apomorphies. Interrupted line shows a fragment of phylogenetic tree which cannot be reconstructed.
In the cases when phylogeny cannot be reconstructed, and holophyletic taxa characterized by apomorphies cannot be established, we can established temporary taxa characterized by plesiomorphies only – plesiomorphons (see I.5.3.2). Such taxon for which autapomorphies are unknown can appear to be paraphyletic or holophyletic. If subsequently paraphyly of this taxon is proved, it is broken, if its holophyly is proved, this taxon is retained. Such approach agrees with the cladistic principle of systematics.
Apologists of the traditional systematics do the same in their practices: they accept existence of a paraphyletic taxon, but do it only until its paraphyly is not proved. At the same time traditionalists declare that both holophyletic and paraphyletic taxa should exist – i.e. both synapomorphies and symplesiomorphies are natural characters and thus are good enough to unite supra-species taxa on bases of them. Traditionalists declare that the cladistic systematics is too poor because it reflects only cladogenesis but does not reflect anagenesis (which is termed also «phyletic evolution», «degree of divergence» or «temps of evolution»). However, we can know out about existence of the cladogenesis (i.e. tree furcation) only in the case if it is supplied with anagenesis (i.e. with getting apomorphies), because only the cladistic analysis (i.e. analysis of apomorphies) allows to clarify furcations of the phylogenetic tree. Thus, the statement that the cladistic systematics does not reflect anagenesis, is wrong; in the cladistic systematics the anagenesis is reflected in the same degree as in the traditionalistic systematics.
Often we can here that anagenesis can have different degree, and that traditionalists, in contrast to cladists, take into account the degree of anagenesis when build classification. However, there is no any distinct definition how to estimate the degree of anagenesis, and the same evolutionary transformation with equal reason can be estimated as a very large or a very small anagenesis.
For example, the taxon winged insects (Pterygota) differs from primarily wingless insects not only by presence of complicatedly organized wings, but also by modification of the whole structure of wing-bearing thoracic segments (see Chapter VII: Pterygota); because of this we could say that origination of this taxon of high rank was accompanied by a large anagenesis (or aromorphosis, if use the terminology by A.N. Severtzov). But in this case it becomes unclear how large should be regarded that anageneses in result of which numerous secondarily wingless species and infra-species forms originated among Pterygota, because in some of these forms not only wings are lost, but the thorax is transformed in such a manner that lost all features characteristic of Pterygota.
In order to estimate degree of anagenetic component, traditional systematics uses a subjective category character weight, but the principle of character weight is not explained. In fact, usually the largest weight is attributed to that character, which represents the oldest apomorphy of the largest group.
For example, if it is possible to chose between the classifications I and II, that classification is chosen, in which to the taxon of highest rank corresponds the oldest apomorphy.

In this case, in the classification I the taxon of highest rank is Pterygota, which have a set of autapomorphies «1» – presence of wings and others [see Chapter VII: Pterygota (1)–(6)]. In the classification II the taxon of highest rank is Amyocerata, which have an autapomorphy «3» – reduction of muscles in antenna flagellum [see Chapeter VI: Amyocerata (1)]. Majority of systematicists, including the authors who name themselves traditionalists, chose the classification II, where the oldest know autapomorphy «3» corresponds to the taxon of highest rank Amyocerata. This is done in spite of the fact that this character (simplification of antenna structure) is poorly visible and has no any positive significance for insect life. Accepting the classification II, they reject the classification I, in which the taxon of highest rank Pterygota has younger apomorphy «1»; this is done in spite of the fact that this character (appearing of wings) played a colossal role in the phylogenesis of insects.
Here, as in all other cases, weight of character is determined not by properties of the character itself, but by position of the character of the cladogram. Thus, the concept «degree of anagenesis» represents only a derivative of the concept «cladogenesis», but not a separate objective reality.
Some authors try to estimate degree of anagenetic component by counting characters; these attempts are evidently meaningless, because morphological characters are non-discrete and can not be counted (see above, I.4.2.6.3). Theoretically, the degree of anagenesis could be objectively estimated if count the number of unique nucleotide changes, but at the present we have no such data.
Our knowledge about phylogeny always changes while new facts are accumulated.
In the example discussed here, the complex of characters «1» was not described enough and was not used in classifications in XVIII – beginning of XIX century (particularly, at that time it was unclear which insects have no wings initially and which have lost them secondarily, so wingless Parasita Latreille 1796 were placed separately from winged and some secondarily wingless insects attributed to Pterodicera Latreille 1802). Later (in the end of the XIX century) the characters of complex «1» became better investigated, and this allowed to reconstruct the phylogenetic branch Pterygota; structure of gnathal pouches of Entognatha (character «2») was also investigated, which allowed to outline this taxon; but phylogenetic relations between Pterygota, Entognatha and Triplura (named at that time Ectotropha, Ectognatha, or «Thysanura» s.str.) remained to be unclear. Basing on such knowledge about phylogeny, the taxon Hexapoda was divided into two subordinate taxa – a plesiomorphon Apterygota and a holophyletic taxon Pterygota.
Still later (Imms 1938) a distribution of the character «3» among arthropods was discovered, that allowed to reconstruct phylogenetic tree more exactly and subsequently establish the taxon Amyocerata Remington 1954 (or «Ectognatha» sensu Hennig 1953). On this base the plesiomorphon Apterygota was broken, and within Hexapoda two holophyletic taxa were established – Entognatha and Amyocerata; the formerly established holophyletic taxon Pterygota is retained, becoming a subordinate taxon in Amyocerata.
Other illustrations of the rule of changing the classification in the direction of the cladistic ideal as the features are studied, are the classifications discussed below, in Chapters IV–X (see Euarthropoda: «Classifications» I–II, Pterygota: «Classifications» I–VIII). In an abstractive form this rule is illustrated in Fig. 1.7.
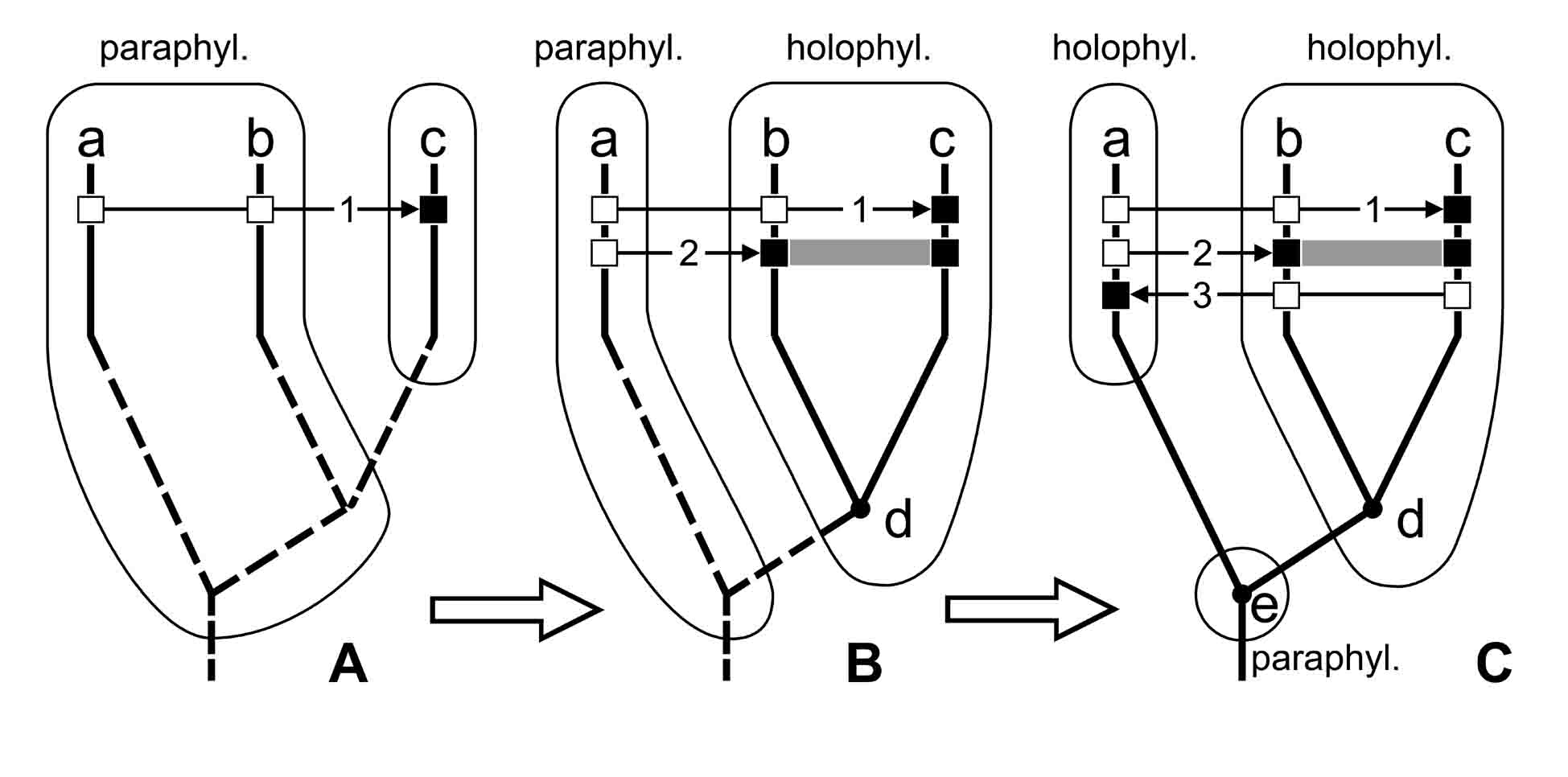
Fig. 1.7. Change of classification in course of discovering of new apomorphies and reconstructing phylogeny. Abbreviations: a–e – taxa (a–c – existent, d–e – ancestral and, as usual, hypothetic); phylogenetic tree is shown by thick line (integral line – reconstructed portions, interrupted line – unknown portions); taxa are outlined by thin line (integral line – known ones, interrupted line – hypothetical ones); 1, 2, 3 – number of characters; thin arrow shows direction from plesiomorphy (white square) to aopomorphy (black square); holophyl. – holophyletic taxon; paraphyl. – paraphyletic taxon; polyphyl. – polyphyletic taxon; thick arrows show change of classificatiom built for the same organisms.
Here the paraphyletic taxon a-b, as our knowledge about previously unknown apomorphies 2 and 3 accumulats, disbands with the formation of a smaller paraphyletic taxon (taxon a in Fig. 1.7B), and than (Fig. 1.7C) is substituted by a holophyletic taxon, which is characterized by an apomorphy 3 and does not include a hypothetical ancestral species e. While knowledge is accumulated, classification approximates to such condition when paraphyletic taxon is restricted to one species which is a common ancestor for two (or more) other taxa of the same rank (this can be the paraphyletic taxon e in Fig. 1.7C). Since this ancestor is usually unknown and is only a hypothetical one, a formal taxon for it is not established, because of this the whole classification consists of holophyletic taxa only. Such classification satisfies completely not only apologists of traditionalism, but apologists of cladism as well (because paraphyletic taxa are formally absent there).
The cladistic principle can be regarded not as a principle according to which classification should be built just now, but as an ideal to which classification should approximate during its change as our knowledge about organisms is accumulated.
Changes of classification make it closer to the cladistic ideal, but this ideal can be reached only when the phylogeny will be completely reconstructed. The phylogeny can be completely reconstructed only when cladistic analysis of all characters will be done. In order to do this, it is necessary to investigate all properties of organisms, i.e. to answer completely all questions posed to biology, and thus close this science. Since this is not expected in the foreseeable period of human history, the classification of living organisms will change always, approximating to the cladistic ideal and never reaching it.
Unfortunately, this makes classification non-stable and creates many inconveniences for investigators. Non-stability of the recent classification is inevitable, because the classification should be natural, and natural features of living organisms on which it is based are in many respects unknown and are intensively studied. If refuse from natural classification and create an artificial one, it would be non-stable because of its subjectivity.
I.5.3.4. Reasons of disagreements between cladists and traditionalists
Everything said in the previous division can provoke an impression that disagreements between cladists and traditionalists about concrete classifications should not be present at all, and the whole controversy could be restricted to explanation of systematics principles only. However, actually disagreements do appear on concrete classifications of this or that group of organisms. Real reason of these disagreements lies not in difference between principles of cladism and traditionalism, but in different opinions in which degree the classification should be stable or non-stable.
As it was shown above, natural classification of animals can not be stable in principle, and should be changed when opinion about phylogeny changes. However, many investigators are prone to demonstrate a justified conservatism, trying not to bring changes to classification until necessity of these changes becomes evident. Apologists of the traditionalistic systematics are prone to demonstrate greater conservatism in this respect, trying to preserve paraphyletic taxa as long as possible. In contrast to them, some apologists of the cladistic systematics demonstrate extremism, trying to change classification immediately when a new theory about phylogeny appears. The reason is that according to the cladistic principles, classification should be built as a cladistic one just now (while actually it can only approximate to this condition – see above). Sometimes a cladist, being attended by a tusk to build cladistic classification immediately, and at the same time not having enough information for phylogeny reconstruction, suggests a non-grounded phylogenetic hypothesis and changes classification on its basis.
If this non-grounded classification can be disproved, opponents do this and thus the argumentation concentrates around concrete questions about phylogeny. However, the a cladist can suggest such evidently wrong phylogenetic hypothesis, which cannot be quickly disproved. This can be done by one of two ways. The first way is to «reconstruct» the phylogeny by such computer method which is not understood by anybody and is grounded on nothing (see I.4.2.6 above), so that even the author of this «phylogeny» can not explain what are his arguments and refers only to a computer program written by somebody; because of this his arguments cannot be disproved. In this case some traditionalists, having no possibility to hold scientific discussion, and at the same time having no possibility to agree with the suggested change of classification, explain their position as a disagreement between cladism and traditionalism.
The second way to provoke disagreements is the following: a cladist who wants to reconstruct phylogeny by temps speeded up, investigates for the first time some difficultly studied character (in contrast to shuffling characters with help of computer, this mode of business is useful itself and in all cases brings a positive contribution to science). For investigation of this new character a lot of forces, time and means are spent, as a result of which this character becomes investigated for a very little number of representatives of very large taxa. After it, this character is extrapolated to all non-investigated members of these taxa, on base of which a cladogram is built and classification is changed. In order to disprove this phylogeny reasonably, it is necessary to study the same character in other representatives of the discussed taxa, but opponents have no forces, time and means for this work; at the same, dubiousness of the suggested phylogeny does not allow to agree with it.
Let us imagine what would happened if people had no colour sight and would have to determine colours with help of difficultly accessible expansive analysers. In this case some investigator who got a rare possibility to work with such analyser, would found out that the housefly has grey colour, while the earth-boring scarab beetle and the beautiful demoiselle have blue colour which is rare in nature; if the blue colour is regarded as a synapomorphy, in this case, taking into account that nothing is known about colour of other insects, beetles and damselflies could be placed to a common taxon opposed to flies.
Some modern phylogenetic hypotheses based on poorly investigated characters look nearly like this: they ere improbable, but at the same time non-disprovable. In such cases some traditionalists also try to explain their negative opinion on the new classification not by disagreements on concrete points of phylogeny, but by disagreements on general principles of systematics.
I.5.4. RELATIVE AND ABSOLUTE RANKS OF TAXA
Above, there were discussed the principles, based of which animal species are grouped in taxa. Now let us discuss the principles on which a form of classification is based. These principles are usually called Linnaean ones, while only some of them were suggested by C. Linnaeus for the first time, and others were taken by him from the works of predecessors. We can call the following principles of the Linnaean classification which distinguish this classification from many others: (1) strict hierarchy of the classification (by other words, presence of relative ranks of taxa); (2) presence of absolute ranks of taxa and (3) inequality of ranks, i.e. presence of basic (obligatory) and non-basic (additional) ranks, and a special status of species and genus. Among these three principles, the first one is grounded by the modern theory about phylogenetic nature of that «Natural System» which the Linnaean classification has to reflect; this hierarchical principle with necessary corrections should be used in the modern systematics as well (see I.5.4.1 below). The two other principles are widely accepted till now, but they do not reflect modern ideas about scientific base of animal classification, and their usage in systematics is non-expedient (see I.5.4.2 and I.5.4.3 below). Because of this, some modern authors try to avoid the absolute ranks. However, until recently, it was not possible to completely dispense with ranks, since the modern rank-based nomenclature of taxa is associated with absolute ranks. Now with help of a new rankink-free nomenclature of supra-species taxa (see I.6.4 below), a possibility appears to write down classification without absolute ranks, not coming to disagreement with the existent rules of the rank-based nomenclature.
I.5.4.1. Strict hierarchy of classification
It is regarded to be generally accepted, that animal classification should be built according to a strict hierarchical principle: each taxon is attributed to a taxon of higher rank, and the same taxon can not be attributed to two or more taxa of equal ranks. In this respect the classification of species of living organisms differs from classifications of many other natural objects (chemical elements, matters, et al.). For example, in the classification of chemical matters, it looks quite normal that the amino acids are attributed to acids, and at the same time they are attributed to organic matters, while neither the organic matters in general are not attributed to acids, nor the acids in general are attributed to organic matters. In contrast to this, in the classification of living organisms, it is completely impossible that, for example, some order would belong to two classes at the same time.
Since old times, the system of living organisms was created not jujst as a convenient classification, but as a reflection of a objectively existing pattern in nature (as wrote C. Linnaeus, «not a character determines a genus, but a genus determines a character»). However, only with development of evolutionary theory, the natural mechanism which provides this regularity, becomes more or less clear. Integrity of each taxon is provided by origination of all members of this taxon from a common ancestor, and a hierarchy of the classification is provided by the fact that increasing of species number takes places by the way of divergence (division of one ancestral species into two or several ones), while origination of one species from two or several ancestral ones is in most cases impossible.
Here we have to pay attention to one detail which is often not taken into account. A hierarchical natural system with more or less distinctly outlined species and taxa of higher ranks, is formed not by a whole phylogenetic tree, but only by its fragments available for our investigation: these are the recent section of the phylogenetic tree (i.e. a totality of all recent organisms) and fossils which are preserved. For these objects available for investigation, we can built a non-conflicting hierarchical classification, where all supra-species taxa are separated by natural breaks. But if take a whole totality of organisms existed in any time (that can be done only mentally), they form an integral phylogenetic tree, i.e. a non-broken chain of generations, which is not divided neither to isolated species, nor, all the more, to taxa of higher rank (Fig. 1.1). If we imagine a classification of all organisms which lived on the Earth, then in this classification the boundaries between taxa will appear to be completely artificial: a boundary between neighbouring taxa of any (even a highest) rank in a certain place will obligatorily cross a chain of generations in such a manner that a mother will be placed in one taxon and her own child in another. C. Linnaeus in his «Philosophy of botany» wrote: «Artificial classes substitute natural ones, while all natural classes are not discovered: when with discovering of many new genera they will be founded out, it will be quite difficult to determine distinct boundaries of classes».
I.5.4.2. Absolute ranks
In the Linaean system, besides the relative ranks (see above), there are absolute ranks (such as class, order, genus, species, et al.). In accordance with this, in the same classification, not only groups uniting different sets of species, but also groups identical in composition but differing in absolute ranks, are considered as different taxa.
For example, if an order Protephemeroidea includes a single family Triplosobidae, where is included a single genus Triplosoba with a single species Triplosoba pulchella, the taxa Protephemeroidea, Triplosobidae, Triplosoba and T. pulchella are identical from biological point of view, but have different ranks and because of this they are formally regarded as four different taxa.
If a taxon unites several taxa of the next rank, it is termed polytypic. If a taxon includes only a single directly subordinated taxon, it is termed monotypic. From its directly subordinated taxon, the monotypic taxon differs by nothing except for an absolute rank, i.e. it has no own biological meaning. The concept of monotypy is purely formal: for example, if several species are united into one family and one genus, this family would be regarded as monotypic; but if the same family with the same set of species is divided to several genera, the family would be regarded as polytypic.
Generally accepted ranks used in zoology are listed in Tables I.3 and I.4.
Table I.3. Ranks of zoological taxa in decreasing sequence
|
Single |
Plural |
Abbreviation |
Satatus |
Source |
|
imperium |
imperia |
imp. |
additional |
Linnaeus
1758 |
|
regnum |
regna |
regn. |
main |
Linnaeus
1758 |
|
phylum |
phyla |
phyl. |
main |
Haeckel 1866
(phylum) |
|
classis |
classes |
cl. |
main |
Linnaeus
1758 |
|
legio |
legiones |
leg. |
additional |
Latreille
1806 |
|
cohors |
cohortes |
coh. |
additional |
Latreille
1806 |
|
ordo |
ordines |
ord. |
main |
Linnaeus
1758 |
|
familia |
familiae |
fam. |
main |
Latreille
1802 (formerly
used instead of order) |
|
tribus |
tribus |
tr. |
main |
Cuvier 1817 (formerly
used for ranks either higher than family, or lower than genus) |
|
genus |
genera |
gen. |
main |
Linnaeus
1758 |
|
species |
species |
sp. |
main |
Linnaeus
1758 |
|
varietas |
varietates |
var. |
additional |
|
|
forma |
formae |
f. |
additional |
|
|
Prefix |
Comment |
|
super- |
cannot be used for genus |
|
sub-
|
can be used for all ranks |
|
infra- |
in zoology is not used for genus and species |
|
subter- |
in
zoology is not used for genus and species |
Among the ranks listed in Table I.3, regnum (kingdom), classis (class), ordo (order), genus (genus) and species (species) were used as obligatory ones in the classical works by C. Linnaeus (1736–1796). Later, as obligatory ranks, familia (family) and phylum (phylum) (which sometimes was also called typus) were added.
In the 10th edition of the book «Systema Naturae» (Linnaeus 1758), which recently is officially accepted as the starting point of zoological nomenclature (see I.6 below), classification looks as the following. Empire Naturae (nature) is divided to 3 kingdoms – Animale (animals), Vegetabile (plants) and Lapideum (minerals). Each kingdom is divided into classes, particularly the kingdom Animale is divided into 6 classes – Mammalia (mammals), Aves (birds), Amphibia (composite group of vertebrates), Pisces (fishes), Insecta (arthropods) and Vermes (other animals). Each class is divided into orders, particularly the class Insecta is divided into 7 orders – Coleoptera, Hemiptera, Lepidoptera, Neuroptera, Hymenoptera, Diptera and Aptera. Each order is divided into genera. The order Eleuterata is divided into 25 genera; among them, besides 22 genera recently attributed to beetles, there are genera Forficula (earwigs), Blatta (cockroaches) and Gryllus (orthopterans in wide sense). The order Hemiptera (corresponding to a modern taxon Condylognatha) is divided into 7 genera. The order Lepidoptera (lepidopterans in modern sense) is divided into 3 genera. The order Neuroptera (corresponding to nothing in modern classifications) is divided into 6 genera – Libellula (odonates), Ephemera (mayflies), Phryganea (caddisflies and stoneflies), Hemerobius (neuropteroids, copeognathans and winged specimens of termites), Panorpa (scorpion-flies) and Raphidia (snake-flies). The order Hymenoptera (hymenopterans in modern sense) is divided into 8 genera. The order Diptera (dipterans in modern sense) is divided into 10 genera. The order Aptera (corresponding to nothing in modern classifications) is divided into 14 genera – Lepisma (triplurans), Podura (springtails), Termes (wingless specimens of termites), Pediculus (lice), Pulex (fleas), Acarus (mites), Phalangium (opilions, thelyfons and phryns), Aranea (spiders), Scorpio (scorpions), Cancer (crabs), Monoculus (some crustaceans et al.), Oniscus (oniscides et al.), Scolopendra (centipedes et al.) and Julus (millipedes). Each genus is divided into species. In some orders genera are grouped into taxa of intermediate ranks. Particularly, in the order Aptera the first five genera are united into a group without rank and without name – this group is characterized by six legs and distinct head; the next six genera are united into a group which is characterized by many legs and indistinct head; and the last three genera – into a group characterized by many legs and distinct head. Some genera are subdivided into intermediate taxa, each of them is divided into species. Particularly, the genus Gryllus is subdivided into taxa without ranks Mantis (raptorial mantids and stick-insects), Acrida, Bulla, Acheta, Tettigonia and Locusta (the last five ones correspond to saltatorial orthopterans).
Subsequently, not only the hierarchy of taxa was changed approaching to the natural one (about the principle of this changing see above), but the absolute ranks changed as well. Some of the Linnaean taxa which are retained now, have retained their ranks without change up to our days: these are, for example the classes Mammalia and Aves, the orders Lepidoptera, Hymenoptera, Diptera. In other taxa retained till our days, ranks are greatly changed: for example, in Forficula, Libellula, Ephemera, Panorpa, Raphidia, Lepisma, Podura, Pediculus and Pulex ranks have grown from genus to order or higher; in Scolopendra and Julus ranks have grown from genus to class; in some taxa of Vermes ranks have grown from genus to phylum.
Ranks represent a usual component of systematics; morever, in accordance with the International Code of Zoological Nomenclature (see I.6 below) those taxa which fall under the rules of this Code can only be assigned official names if these taxa are assigned concrete ranks.
However, expediency of ranks using is doubtful. Only one rank – species – has a scientifically grounded definition: the criterion of biological species is the presence of reproductive isolation from other species and absence of reproductive isolation within the species. The significance of all other ranks is reduced only to showing of hierarchical subordination of taxa: a taxon of higher rank is divided into taxa of lower ranks. Some authors believe that ranks have to bear some more scientific information, while they disagree on which one. In accordance with meaning they want to attribute to ranks, they suggest rank criterions conflicting one with another: some authors suggest to connect ranks with degree of divergence of taxa, and others – with age of taxa.
It is impossible to connect ranks with degree of divergence, because there is no universal criterion for estimation of the divergence degree.
The author of phylogenetic systematics W. Hennig regarded that since in the phylogenetic systematics the relative ranks of taxa (i.e. their position in hierarchy in relation one to another) depend upon the relative time of divergence (i.e. upon succession of branching of phylogenetic tree – see I.5.2.1), then the absolute ranks of taxa (i.e. phyla, classes, orders, and so on) should depend upon the absolute (i.e. geological) time of divergence. In order for the ranks of insect taxa established by geological age, to be most consistent with the traditionally accepted ranks of these taxa, Hennig proposed the following correspondence between the ranks and the time of separation of taxa: class – from Cambrian to beginning of Devonian; order – from end of Devinian to end of Permian; family – from beginning of Triassic to end of Early Cretaceous; tribe – from beginning of Late Cretaceous to end of Oligocene. However, if accept this scale for taxa of vertebrates, they should be lowered in their ranks; particularly Mammalia should be regarded not as a class but as an order; some other taxa should be raised in their ranks: particularly some taxa known beginning from Ordovician and regarded as genera in the class Ostrapoda, should be regarded not as genera but as classes. Usage of this rank criterion meets following difficulties. The absolute time of divergence can be determined only on the base of paleontological data. At the same time, it can be reliably established that the divergence happened no later than such and such a time (if the remains of organisms that arose as a result of this divergence are found in the sediments of this time), but there is no method which would allow one to state that this divergence happened not earlier than a certain time. Because of this, hypotheses about time of divergence often have a very large dispersion. For many groups of organisms there are no fossils at all, and for them this criterion of ranks is not acceptable.
Since there are no generally accepted criteria for supra-species ranks, in recently existent classifications ranks are establishes in arbitrary manner. The ranks are partly natural, partly artificial component of systematics: their relative meanings can be natural, while their absolute meanings are artificial. If classifications differ one from another only by absolute ranks, there is no principle difference between these classifications.
Usage of ranks often causes a wrong idea that any taxa of the same rank have something common one with another. As a result of this, in some scientific works taxa of the same rank are used as comparable elements. For example, faunas are compared by a number of common genera, or phylogeny is discussed on level of families. Actually taxa of the same rank are comparable elements only if these are sister-taxa inside one taxon of higher rank: for example, genera of one family are comparable one with another, while genera from different families only formally are regarded to be taxa of the same rank, and their comparison is groundless.
In the chapters III–X of this book, all taxa are given without absolute ranks, that appeared to be possible because of consistent usage of rank-free nomenclatures (see I.6 below); before characteristic of each taxon, under the heading «Rank», there are listed that ranks which are most often attributed to this taxon.
I.5.4.3. Inequality of absolute ranks
Among absolute ranks, basic and additional ranks are distinguished (see Table I.3). Among the basic ranks (species, genus, family, order, class, phylum, and kingdom), the ranks species and genus have special status. The basic ranks are mandatory, while the additional ranks are optional.
If in a classification number of hierarchical levels is less than number of existent basic ranks, then all basic ranks are used, even if this is not necessary.
For example, if a single species Triplosoba pulchella cannot be placed in any previously established order, for this species a special monotypic order Protephemeroidea is established; in this order a new monotypic family Triplosobidae and a new monotypic genus Triplosoba are established (both have circumscription and diagnosis coinciding with the circumscription and diagnosis of the species and the order).
In such manner, the classification has a surplus of taxa and names.
If in a classification the number of hierarchical levels is more than number of existent basic, then all the basic ranks and the required number of additional ranks are used. With this, attributing of a basic or an additional rank to this or that taxon is arbitrary.
As an example, let's take a classification of recent groups of arthropods written without ranks and than attribute ranks to these groups by two different ways (in the modern literature much more manners of attributing ranks for these taxa are found):

Since the additional ranks are non-obligatory ones, many authors omit them, and together with them omit taxa to which these ranks are attributed, and omit diagnoses of these taxa. As a result, in some publications the phylum Arthropoda contains the class Hexapoda, for which a long characteristics is given, but at the same time nothing is said about the taxon Amyocerata; in other papers, vice versa, a long characteristics of the class Amyocerata is given, while the taxon Hexapoda is ignored. This causes a wrong impression that we speak about two basically different classifications. A curiosity is, that some people seriously think that somebody suppressed one of these taxa, regarding it to be an artificial group; usage of the rank-based nomenclature faciliates to appearance of such misunderstanding (see I.6.6.2 below).
I.5.4.4. Problem of genus
The special status of the rank of the genus is expressed in the fact that, according to the Linnaean principle, enshrined in the International Code of Zoological Nomenclature (see below, I.6.3), a species can receive a binary name only if it is at least formally assigned to some genus. Because of this, in order to name a species, it is necessary not only to place it into some supra-species taxon, but also chose which of the supra-species taxa including this species should be regarded as a genus. When the generic rank is moved from one supra-species taxon to another, species names are changed (while when ranks of other supra-species taxa are changed, only names of these taxa are changed, but not names of species included in them). At the same time, the generic rank is the less stable among others, being most often moved from one systematic taxon to another.
This contradiction is caused by the fact that during the development of taxonomy, the same word «genus», was understood as three fundamentally different things. It is necessary to distinguish between the Aristotelian genus, the Linnean genus and the Latrailean genus.
The Aristotelian genus is a relative concept: it is any category that unites species, while the Aristotelian species is any category in the composition of the genus. According to Aristotle, species are united into genera; if these genera, in turn, are united into a genus of a higher order, then they are species in relation to this superior genus; accordingly, a species can be subdivided into species of a lower order, and then it is a genus in relation to them. Thus, the entire hierarchy is described by two categories – species and genus. Aristotle believed that hierarchy was a fundamental property of all nature, so the Aristotle's species and genus were fundamental categories.
The Linnaeus genus, in contrast to the Aristotelian genus, is not relative, but absolute. Linnaeus called genera are not any taxa that unite species, but taxa of only one hierarchical level: for each species, only one higher taxon is called a genus, and a taxon that unites several genera is called not a genus of a higher order, but otherwise (for example, an order). Thus, the hierarchy is described not by two relative categories (species and genus), but by several absolute categories – species, genus, order, class, etc. In this system, the genus appeared to be not a fundamental category, but only one of several equivalent ones. In spite of the fact that Linnaeus changed the meaning of the words «species» and «genus», he continued to consider species and genus as fundamental categories and regarded the species and genera to be more natural than taxa of other ranks. Linnaeus saw the peculiar status of the species in that intraspecific taxa evolve, while species and all taxa above the species are unchanged; at present, we consider this idea erroneous, but if the term «species» is applied to a biological species (see I.3)], then it really has a peculiar status, because intraspecific taxa do not have reproductive isolation, while species and all taxa above the species are reproductively isolated one from another. As for the peculiar status of the genus, Linnaeus did not explain it in any way; apparently, the idea of the fundamental genus concept was simply mechanically transferred from Aristotle's philosophy to systematics.
The binary nomenclature was elaborated by Linnaeus proceeding from the fact that initially genera were the most stable taxa in his system. If a natural group was regarded as a genus, that genus, with its diagnosis and circumscription (i.e. boundaries) was retained in subsequent changes of the classification. If new species were discovered that corresponded to the diagnosis of a previously established genus, they were placed to this genus. Thus, number of species in the genus increased, but the genus remained to be the same. When number of species increases, it becomes necessary to bring them in order; for this purpose number of ranks and taxa should be increased. Linnaeus introduced additional ranks placing them, when necessary, both above the genus and below the genus. Thanks to this, increasing of species number did not prevent stability of genera and did not lead to changing of names for species previously described. Only in the case if an artificial group was established as a genus, such genus was disbanded.
In the Linnaean system, only taxa of basic ranks (class, order, genus and species) had uniform names, while for taxa of additional rank, no distinct principles of nomenclature existed: for example, the genus Gryllus was divided into taxa without ranks with names in singular – Mantis, Acrida, Bulla, Acheta, Tettigonia and Locusta, but the Cicada was divided into taxa without ranks with names in plural – Noctilucae, Foliaceae, Cruciatae, Manniferae, Spumantes and Diflexae.
The Latrailean genus is absolute only in a certain classification, but when the classification is changed, it turns out to be relative. In a concrete classification, for each species, only one concrete superior taxon is called a genus (as in the case of the Linnean genus), but in different classifications the rank of the genus can be assigned to different taxa that are superior to this species. Since the number of known species increases, the rank of the genus is assigned to a taxon of an increasingly lower level. Cirmscription of the Linnaeus genus can be changed only if an error was made when it was establishment; a change in the circumscription of the Latrailean genus is a normal action, not associated with errors. This situation has appeared for the following reasons.
Linnaeus united genera of animals and plants to orders, for which he gave non-typified names. Botanists of that time termed a taxon between genus and classis either an order (Jussieu 1789), or a family (Adanson 1763), and gave typified names for some of these taxa.
Latreille (1802) was one of the first who used in zoology family rank along with ordinal rank, placing family between genus and order in his classification of arthropods; at the same time he introduced additional ranks, both above and below the family. Initially family names were both typified and non-typified, and the typified ones were not unified. Subsequently, systematicists began to form names of all taxa of the family-group from names of type genera, adding to them standardized endings corresponding to ranks – i.e. created typified unified names. Hence, there appeared a convenient method for adding ranks necessary to classify increasing number of newly discovered species, and this method can be applied to taxa above genus only.
There is no convenient method for adding ranks below genus. In zoology, between genus and species only one rank can be added – subgenus (in botany several infrageneric ranks are acceptable).
Usage of the subgeneric rank has its disadvantages, which are why many zoologists avoid it: the subgeneric name does not differ from the generic one, so in order to distinguish them, the rank should be indicated; in the binomen subgeneric name is written in brackets between the generic and specific names, that makes binomen more cumbersome.
As a result of this, when in some genus number of known species increases in such degree that it becomes necessary to introduce a new rank, the new rank is placed not within the genus, but above the genus. This leads to an inflation of genera: while species number increases, generic rank each time moves to a subordinate taxon, thus number of genera grows quickly, and the genus always remains to be a smallest supra-species taxon. So, if a genus was previously was established that satisfiers all requirements of taxonomy (holophyletic, with a clear diagnosis), than later it is divided into several genera not because there were any doubts about its naturalness, but only because the number of distinguishable species has increased.
Inconveniences connected with the inequality of ranks and the inflation of genera, are taken away if use the rank-free principle of systematics and rank-free nomenclature, as it is done in this book (see I.6.4, I.6.5, I.6.7 and Chapters III–X below).