CLADOENDESIS OF EPHEMEROPTERA
|
CLADOENDESIS OF EPHEMEROPTERA |
||||
| zzz | ||||
|
|
|
Pseudopannota/g2
(Panephemeroptera![]() Euephemeroptera
Euephemeroptera![]() Euplectoptera
Euplectoptera![]() Anteritorna
Anteritorna![]() pm.Tridentiseta
pm.Tridentiseta![]()
Tetramerotarsata![]() Liberevenata
Liberevenata![]() Turbanoculata
Turbanoculata![]()
Anteropatellata![]() Baetovectata
Baetovectata![]() Baetungulata
Baetungulata![]()
Labiobaetis/f1=Pseudopannota/g1![]() - Pseudopannota/g2)
- Pseudopannota/g2)
Nomen hierarchicum:
![]() Pseudopannota/g2
[g:1987] (sine Labiobaetis;
incl. Ophelmatostoma, Hemipannota).
Pseudopannota/g2
[g:1987] (sine Labiobaetis;
incl. Ophelmatostoma, Hemipannota).
In circumscription fits:
— gen. Pseudopannota: Kluge & Novikova 2016: 5.
— Pseudopannota/g1: Kluge & Novikova 2016: 5.
Nominal taxa included:
![]() Ophelmatostoma/g [g:1987]
Ophelmatostoma/g [g:1987]
![]() Hemipannota/g [g:1990]
Hemipannota/g [g:1990]
References.
Kluge & Novikova 2016: 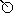

|
Autapomorphies of Pseudopannota/g2 (characters common with Guajirolus/g1 marked by brown). (1) Modification of labrum. Labrum has the following peculiar structure (Kluge & Novikova 2016: Fig.37, 38–39, 74–76). Distal margin lost sclerotized seta-bearing band, and outer side lost constant pair of submedian setae and paired latero-distal setae, which are initially characteristic for Baetidae [see Tetramerotarsata (6) and Turbanoculata (4)]. Instead of this, distal margin is formed by a soft area with membranous cuticle covered by numerous irregularly arranged fine colorless setae (Kluge & Novikova 2016: Fig.74–76); inner side near anterior margin bears a pair of transverse rows of stout pointed spine-like setae, accompanied by a pair of transverse sclerotized ridges; median end of right ridge asymmetrically continues as longitudinal ridge running in proximal direction (Kluge & Novikova 2016: Fig.37, 74, 76). I was unable to find correlation between this asymmetric element of labrum structure with asymmetric elements of adjacent mouthparts – mandibles and hypopharynx. (2) Reduction of mandibular incisors. On both left and right mandibles, incisors are more or less reduced, so that the most projected apical denticle of mandible belongs to kinetodontium (Kluge & Novikova 2016: Fig.6–7, 40–41, 77–79, 92–94). This fact becomes clear if compare the mandible usual for plesiomorphon Labiobaetis/f2=g1 and other Turbanoculata (Kluge & Novikova 2016: Fig.4–5) with mandible of a primitive representative of Pseudopannota/g2, where two denticles of incisor are retained (Kluge & Novikova 2016: Fig.6) and with mandible of a derived representative of Pseudopannota/g2, where incisor is reduced to a single small denticle (Kluge & Novikova 2016: Fig.7). (3) Protuberance on right mandible. Right prostheca is attached in a cavity, just proximad of which a sclerotized protuberance is located (Kluge & Novikova 2016: Fig.6–7, 42, 78–79, 93). (4) Shape of maxilla. Maxilla is different from maxilla of «Baetis-type» (which is characteristic for Labiobaetis/f2=g1 and most other Baetungulata) and is similar to maxilla of «Cloeon-type» (Crass 1947: 64): its biting margin is long, with all three canines and all three dentisetae long, pointed and thin-walled (Kluge & Novikova 2016: Fig.9,12,15, 36, 70). Change between «Baetis-type» and «Cloeon-type» in both directions repeatedly took place in evolution of Turbanoculata. (5) Modification of apical setae of maxilla. Apex of maxilla just laterad of canines bears long setae, which form a regular transverse row, with bases of setae pressed one to another; this row locates either on ventral-apical side only (Kluge & Novikova 2016: Fig.36), or surrounds bases of canines from ventral, apical and dorsal sides (Kluge & Novikova 2016: Fig.70). (6) Pseudo-three-segmented maxillary palp. Maxillary palp [2-segmented – see Baetungulata (3)] looks as thee-segmented, because its first primary segment is divided into two subsegments movably connected one with another; muscle, running from base of maxillary palp to base of the primary second segment, passes through both subsegments of the primary first segment (Kluge & Novikova 2016: Fig.36) (see Index of characters [1.1.42]). Among Pseudopannota/g2, only in camerunense [Pseudocloeon] maxillary palp is vestigial, very thin, lost intrinsic muscle and lost division of the first segment (see below). Initial for Ephemeroptera is three-segmented maxillary palp with muscle present in the first segment only (instead of initial for insects 5-segmented maxillary palp with muscles in four segments). Such 3-segmented palp is retained in a part of Baetidae. As its 2nd segment always lacks intrinsic muscle, articulation between 2nd and 3rd segments does not provide active movements; in several non-related mayfly taxa this articulation undergoes reduction, so that palp becomes 2-segmented. Particularly, such 2-segmented maxillary palp is characteristic for the taxon Baetungulata, to which Pseudopannota/g2 belong (Kluge & Novikova 2011). Maxillary palp of Pseudopannota/g2 is not true 3-segmented, but pseudo-3-segmented, because its first two segments originated from the initial first segment and both contain its muscle. The 2-segmented maxillary palp of plesiomorphon Labiobaetis/f2=g1 has cuticle of both segments entirely membranous, with numerous campaniform sensilla (looking as small rings) located on the first segment (Kluge & Novikova 2016: Fig.8). The pseudo-3-segmented maxillary palp of examined species of Pseudopannota/g2 has cuticle partly sclerotized: it is sclerotized at least on outer side of 2nd segment and on outer side of the distal subsegment of 1st segment. The proximal subsegment of 1st segment has cuticle either entirely membranous, or membranous at least on inner side; campaniform sensilla are concentrated on membranous cuticle of its inner side (Kluge & Novikova 2016: Fig.14; Gattolliat 2002: Fig.1d). In fusca [Pseudopannota] proximal subsegment has sclerotized cuticle on outer side (Kluge & Novikova 2016: Fig.9); on shed exuviae it keeps its form, and only its base is inserted into stipes because of inversion of membranous cuticle of palpiger (ibid. Fig.10). The same in pannota [Pseudopannota] (Kluge & Novikova 2016: Fig.14). In the related species Pseudopannota sp. U, proximal subsegment has cuticle entirely membranous, so that on shed exuviae it is completely inverted and inserted into stipes (Kluge & Novikova 2016: Fig.15). In camerunense [Pseudocloeon] both labial and maxillary palps are weakened and narrowed, so that maxillary palp lost its secondary articulation between subsegments of the first segment. Two parts of the first segment, corresponding to two subsegments, retain difference: distal part retains sclerotization of outer side, and proximal part retains entirely membranous cuticle and campaniform sensilla on its inner side (Kluge & Novikova 2016: Fig.11–13, 70–71). Besides this, in penultimolarva the proximal part is thicker than the distal part (Kluge & Novikova 2016: Fig.11, 71). Because of this, on shed exuviae of penultimolarva, the proximal part of first segment is completely inverted and inserted into stipes (Kluge & Novikova 2016: Fig.12), while on shed exuviae of ultimolarva maxillary palp keeps its form, and only its base is inserted into stipes because of inversion of membranous cuticle of palpiger (ibid. Fig.13). These differences between proximal and distal parts of the first segment testify about their origin from the proximal and distal subsegments, which are expressed in other species of Pseudopannota/g2. Near its apex, second segment of maxillary palp has incision on inner side, which separates a small apical portion (Kluge & Novikova 2016: Fig.9–15, 70–71). Similar incision is present in certain species of Labiobaetis/f2=g1 (Kluge & Novikova 2016: Fig.123), but not in other Baetungulata. Possibly, the apical portion separated by this incision, is a remnant of the primary third segment. (7) Modification of labium. Glossae and paraglossae of labium are more or less elongated, with apical setae elongated, situated either irregularly (Kluge & Novikova 2016: Fig.34, 96), or forming ring-like row (Kluge & Novikova 2016: Fig.68–69). (8) Two-segmented labial palp. The 2nd and 3rd segments of labial palp are completely fused together, and intrinsic muscle of 2nd segment is completely lost (Kluge & Novikova 2016: Fig. 35, 68, 96). The same in some other taxa, but not in plesiomorphon Labiobaetis/f2=g1, whose intrinsic muscle of 2nd segment is always retained (Kluge & Novikova 2016: Fig.126). (9) Pannote condition. Fore protoptera [see Turbanoculata (8)] are widened in such a manner that their bases are brought together (Kluge & Novikova 2016: Fig.18–20); in certain species fore protoptera are fused together at a long distance (ibid. Fig.17). Unlike Pseudopannota/g2, in other Turbanoculata fore protoptera are separated by wider distance (Kluge & Novikova 2016: Fig.16). (10) Modification of larval coxa. Larval coxa has more or less prominent lobe anteriad of trochanter base; shape of this lobe is species-specific; it is either similar on all legs, or different on legs of different pairs (Kluge & Novikova 2016: Fig.50–52, 82–83, 110–111). (11) Differentiation of larval legs. Larval fore leg differs from hind leg by presence of additional spine-like setae on anterior side of femur; middle leg has intermediate structure (Kluge & Novikova 2016: Fig.50–52, 82–83, 110–111). Variable characters of Pseudopannota/g2. Larval shape varies from the «swimming type» (Kluge & Novikova 2016: Fig.59–60) to «crawling type», i.e. shortened and robust with strengthened legs (Kluge & Novikova 2016: Fig.31). Larval claws either with usual number (6–8) of subequal denticles, or with reduced number (3–4), or with 2 unequal denticles. Larval abdominal terga either smooth with scales in semicircular sockets (e.g., in maculosum [Pseudocloeon], camerunense [Pseudocloeon]), or without scales and with prominent relief (e.g., in pannota [Pseudopannota]), or with both scales and prominent relief (e.g., in P. fusca). Tergalii with or without scales and/or relief similar to that of abdominal terga, with or without additional rib along costal margin. Number of tergalii pairs varies from 7 to 6: tergalii of first pair either as large as others, or diminished, or lost. Subimaginal tarsomeres 1–4 other than on male fore legs, either with pointed microlepides, or with blunt microlepides. Hind wings are lost in all known species [see Turbanoculata (10)]; (non-unique apomorphy – see Index of characters [2.2.59]); vestiges of larval hind protoptera either present (in camerunense [Pseudocloeon]), or absent. Discussion. The most peculiar characters of Pseudopannota/g2 coincide with characters of the Neotropical getaxon Guajirolus/g1: In both taxa labrum has numerous fine setae on outer side, soft distal margin and pair of rows of spine-like setae on inner side (but Guajirolus/g1 has no asymmetric sclerite on inner side of labrum). In both taxa mandibular biting denticles are formed by kinetodontium, while incisor is greatly reduced; prostheca (at least right one) is inserted into incision (but Guajirolus/g1 has no protuberance proximad of this incision). In both taxa maxilla has long biting margin with long and delicious maxillary canines and dentisetae, and distal setae are long (but in Guajirolus/g1 these distal setae don't form a regular row). In both taxa maxillary palp is pseudo-three-segmented, with primary first segment divided into two subsegments. In both taxa glossae and paraglossae are elongated, with elongated apical setae. In both taxa larval fore femur is strengthened and bears additional spine-like setae on anterior side. However, Guajirolus/g1 does not belong to Labiobaetis/f1=Pseudopannota/g1: in contrast to it, Guajirolus/g1 retains sterno-styligeral muscle, has different type of gonostyli folding in mature larva and has different shape of imaginal gonostyli. |
Size. Fore wing length 5–6 mm (see Tetramerotarsata).
Distribution. Afrotropical Region.
Nominal species in Pseudopannota/g2:



 berthrandi Demoulin
1967 [Pseudocloeon]
berthrandi Demoulin
1967 [Pseudocloeon]  — typus nominis Pseudopannota
— typus nominis Pseudopannota ![]()
![]() exuv.,
exuv.,![]() ,
,![]()
![]()
camerunense Ulmer 1920 [Pseudocloeon]
![]()
![]() -
-![]() -
-![]() /
/![]()
![]()
camillae Gattolliat 2002 [Pseudopannota (Hemipannota)]
fusca Kluge
& Novikova 2016 [Pseudopannota]
![]()
![]() -
-![]() /
/![]()
![]()
kimminsi Waltz & McCafferty 1987 [Ophelmatostoma]
— typus nominis
Ophelmatostoma
maculosum Crass 1947 [Pseudocloeon]
— typus nominis Hemipannota
![]()
![]()
![]()
modesta Elouard & Gillies (in Elouard & Gillies & Wuillot)
1990 [Pseudopannota (Hemipannota)]
muganinani Elouard & Gillies (in Elouard & Gillies & Wuillot)
1990 [Pseudopannota (Pseudopannota)]
pannota Kluge
& Novikova 2016 [Pseudopannota]
![]()
![]() -
-![]() -
-![]() /
/![]()
![]()
![]() vinckei Demoulin 1973 [Pseudocloeon]
vinckei Demoulin 1973 [Pseudocloeon]
Examined also:
![]() sp.n. (Tanzania)
sp.n. (Tanzania) ![]()
![]() -
-![]() -
-![]() /
/![]()
![]()
![]() sp.n. (Tanzania)
sp.n. (Tanzania) ![]()
![]() -
-![]() -
-![]() /
/![]()
![]()
See also:
Baetovectata
INCERTAE SEDIS ![]()
Turbanoculata
INCERTAE SEDIS ![]()