КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ

![]()
|
КРАТКАЯ СИСТЕМАТИКА НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
X-1.3.1. Hymenoptera Linnaeus 1758 – перепончатокрылые
(Somobialia
Eucaryota Metazoa Monostomata
Haemataria Peritoneata
Metameria
Gnathopoda Euarthropoda
Mandibulata Atelocerata
Hexapoda Amyocerata
Pterygota
Metapterygota Neoptera
Eumetabola
Metabola Mecopteriformia Hymenoptera)
(рис. 10.7.1–10.7.4)
(рис. 10.7.1,
10.7.2, 10.7.3,
10.7.4)
|
Названия, однозначно соответствующие по циркумскрипции: — Hymenoptera Linnaeus 1758; NomCirc. — Aculeata Scopoli 1763; — Piezata Fabricius 1793; — Phleboptera Clairville 1798; — Hymenopteria Rafinesque 1815; — Solenognatha Spinola 1850; — Metabola-Polynephria Brauer 1885 (non Menognatha-Polynephria Brauer 1885); — Lambentia Haeckel 1889; — Hymenopteroidea Handlirsch 1903; — Hymenopterida Pearce 1936; — Panhymenoptera Crampton 1938. По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родового названия Vespa: Vespoides, Vespida, Vespidea. Иерархическое название: Vespa/f1=Cynips/g1 [f:1781(ord.6); g:L.1758(gen.213)] (incl. Xyela). |
|
Ранг. Обычно отряд, иногда одновременно с этим надотряд.
Характеристика Hymenoptera
Особенности имаго. Мандибулы всегда хорошо развиты и сохраняют исходную грызущую функцию. Другие части ротового аппарата – максиллы, нижняя губа и гипофаринкс – также могут также сохранять строение, свойственное исходному грызущему ротовому аппарату, либо преобразованы и адаптированный для питания нектаром цветков. В последнем случае складывающиеся и выдвигающиеся части лабио-максиллярного комплекса (см. Mecopteriformia) бывают удлинены, так что образуют лижуще-сосущий хоботок, способный выдвигаться и проникать в нектарники цветков; такой хоботок независимо возник в разных таксонах перепончатокрылых.
Переднегрудь имеет характерное строение (рис. 10.7.1): латеральные цервикальные склериты (см. Pterygota) неподвижно слиты с плейритами переднегруди; вся плейро-стернальная часть переднегруди (несущая передние ноги) подвижно сочленена с тергитом переднегруди и со среднегрудью. Тергит переднегруди может сохранять подвижное сочленение со среднегрудью (рис. 10.7.1), либо укорочен и слит со среднегрудью; в обоих случаях слитная плейро-стернально-шейная область переднегруди сохраняет подвижность по отношению к среднегруди.
Крыловой аппарат имеет ярко выраженную переднемоторность: задние крылья сцепляются с передними и движутся пассивно. В связи с этим среднегрудь увеличена, а заднегрудь уменьшена. Крылья имеют типичную для переднемоторных насекомых форму: переднее крыло треугольное, с выраженным торнусом; заднее крыло всегда значительно меньше переднего, его костальный (передний) край равен по длине базиторнальному (заднему) краю переднего крыла; так что когда крылья сцеплены, задний край заднего крыла выглядит продолжением торноапикального (дистального) края переднего крыла. Переднее и заднее крыло сцепляются с помощью крючковидных щетинок (hamuli), образующих правильный продольный ряд на костальном крае заднего крыла и зацепляющихся за подогнутый базиторнальный край переднего крыла. Несмотря на то, что задние крылья всегда уменьшены и движутся пассивно, они почти никогда не утрачиваются, так что перехода к морфологической двукрылости не происходит (в отличие от многих других переднемоторных насекомых).
На крыльях имеется сравнительно небольшое число крепких постоянных жилок, соединенных под разными углами и образующими крупные многоугольные ячейки; при этом бывает трудно понять, какие из сторон этих ячеек образованы отрезками исходных продольных жилок, а какие – поперечными жилками (рис. 10.7.1, 10.7.2A–B). На переднем крыле жилка RA сливается с C несколько дистальнее середины переднего края крыла; в месте этого слияния имеется утолщение – птеростигма (pterostigma). Лишь в редких случаях птеростигма вторично утрачена. Жилка, идущая по переднему краю крыла дистальнее птеростигмы сливается с вершиной RS; дистальнее места их слияния наружный край крыла не обрамлен жилкой, и жилки M и Cu1 оканчиваются не достигая края крыла. От Cu назад отходит короткая жилка Cu2, своей вершиной она сливается с вершиной Pcu; это слияние, пересекающее клавальный желоб, находится в дистальной части крыла вблизи заднего (базиторнального) края, около дистального конца подгиба заднего края, обеспечивающего сцепку крыльев. В случаях крайней редукции жилкования (независимо произошедшей в нескольких неродственных друг другу таксонах) на переднем крыле имеется лишь RA, птеростигма (или краевая жилка на ее месте) и отходящий от нее назад-дистально короткий отрезок RS.
В покое крылья складываются обычно плоско на спине.
У большинства перепончатокрылых крылья развиты; иногда крылья развиты только у самцов, а у самок вестигиальные или отсутствуют; реже крылья отсутствуют у обоих полов; у муравьев (Formicidae) самки бесплодной касты бескрылые, тогда как самки плодовитой касты и самцы крылатые, но самки сбрасывают крылья после оплодотворения.
На передней ноге одна из двух шпор (передняя, если нога направлена в сторону) преобразована в аппарат для чистки антенн – стригил (strigilis); при этом вторая (задняя) шпора может сохраняться, оставаясь неспециализированной, либо утрачена (у перепочатокрылых, объединяемых в таксон Unicalcarida Schulmeister, Wheeler & Carpenter 2002). У некоторых пилильщиков (Tenthredinidae) обе шпоры передней голени имеют неспециализированное строение; вероятно, это вторичная утрата исходной для Hymenoptera специализации. В редких случаях шпоры передней голени утрачены.
Небольшой проксимальный участок бедра отделен от остальной части бедра кольцевым швом и выглядит как второй вертлуг; этот участок называют вертлужок (trochantellus), или префемур (praefemur) (рис. 10.7.1). Внешне кажется, что нога имеет «двучлениковый» вертлуг, хотя на самом теле истинный вертлуг здесь одночлениковый. Такой «двойной» вертлуг выражен на всех или на некоторых ногах многих Symphyta и Parasitica, но в некоторых случаях утрачен; в частности, он отсутствует у всех Monotrocha.
Тергиты прегенитальных сегментов брюшка занимают не только дорсальные, но и латеральные поверхности сегментов; их латеральные лопасти направлены вентрально и облегают латеральные области стернитов (кроме первого сегмента брюшка, стернит которого отсутствует); при этом дыхальца (имеющиеся на I–VIII уромерах или на части из них – см. Amyocerata) находятся на склеротизованных тергитах (рис. 10.7.1).
Гениталии самца у всех Hymenoptera имеют единый сложный план строения и в покое втянуты внутрь брюшка (рис. 10.1.4B и 10.7.2C–D).
У самки обычно сохраняется первичный яйцеклад, состоящий из 3 пар створок (см. Amyocerata); при этом первые и вторые створки (т.е. пара VIII кинетапофизов и слитые между собой IX кинетапофизы) сцеплены олистетерами и образуют режущий или колющий яйцеклад, а третьи створки (стилигеры IX уромера) образуют ножны характерного строения (см. Eumetabola) (рис. 10.7.1). Среди Metabola перепончатокрылые сохраняет наиболее полное строение первичного яйцеклада, тогда как у всех прочих Metabola яйцеклад в той или иной мере упрощен или утрачен. Следует обратить внимание на очень большую консервативность в сохранении первичного яйцеклада у Hymenoptera: у представителей с самыми различными способами откладки яиц яйцеклады по-разному модифицированы и по-разному функционируют, но сохраняют все элементы исходного плана строения, лишь в редких случаях подвергаются редукции. У представителей плезиоморфона Symphyta, личинки которых питаются травами и листьями, яйцеклад приспособлен для пропиливания ткани растения: он недлинный, с пиловидным вентральным краем первых створок; в связи с такой формой и функцией яйцеклада этих насекомых называют пилильщиками. У многих других перепончатокрылых (Xyelidae, Uroceridae, многие Euhymenoptera) яйцеклад прямой, удлиненный, приспособлен для прокалывания. У Euhymenoptera колющий яйцеклад, помимо функции проведения яиц, может использоваться для проведения ядовитого секрета; с помощью такого яйцеклада-жала самка парализует насекомое и откладывает на него яйца. У Euhymenoptera-Monotrocha яйцеклад утратил функцию проведения яиц и сохранил только функцию жала. Лишь у немногих перепончатокрылых яйцеклад утрачен.
Особенности личинок. Если личинка имеет глаза (развивающиеся независимо от имагинальных глаз – см. Metabola (1Ca), то имеется одна пара глаз, каждый из которых образован слиянием омматидиев фасеточного глаза и имеет единую выпуклую кутикулярную линзу (вместо разрозненных стемм, имеющихся у большинства других личинок насекомых с полным превращением). Глаза имеются у личинок многих Symphyta (рис. 10.7.3A), но у Uroceridae и Euhymenoptera (составляющих большинство видов перепончатокрылых) личиночные глаза полностью утрачены в связи со скрытым образом жизни (рис. 10.7.4A). Личинка либо имеет гусеницеобразную форму тела с длинным туловищем, небольшими, ногами [см. Metabola (1Ccα)] и ложноножками на брюшке (у многих Symphyta – рис. 10.7.3A), либо безногая.
Гапло-диплоидное определение пола. Из оплодотворенных яиц развиваются диплоидные самки, а из неоплодотворенных яиц – гаплоидные самцы. Соответственно, только у самок оогенез сопровождается мейозом, тогда как у самцов сперматогенез происходит без мейоза. Благодаря гапло-диплоидному определению пола степень генетического сходства между родными сестрами, получившими половину своих генов от одного и того же гаплоидного самца, составляет 3/4 (тогда как между матерью и дочерью она лишь 1/2); так что естественный отбор может поддерживать такие формы поведения, при которых самка выращивает своих сестер в ущерб собственным дочерям. Благодаря этому в некоторых таксонах жалящих перепончатокрылых, у которых самки выкармливают личинок в гнезде, независимо несколько раз возникали одинаковые формы эусоциальности (см. «Классификации» III: Monotrocha).
Длина тела 0,2–60 мм.
Триас – ныне (рис. 2.2). Наиболее древними известными перепончатокрылыми являются представители современного таксона Archihymenoptera из раннего триаса Средней Азии (Расницын 1969; Rasnitsyn & Quicke 2002).
По разным оценкам от 100 000 до 200 000 видов и более; всесветно.
Классификации Hymenoptera. Традиционно перепончатокрылых делят на плезиоморфон Symphyta и голофилетический таксон Apocrita, который, в свою очередь делят на неоправданно большой плезиоморфон Parasitica и голофилетический таксон Monotrocha.
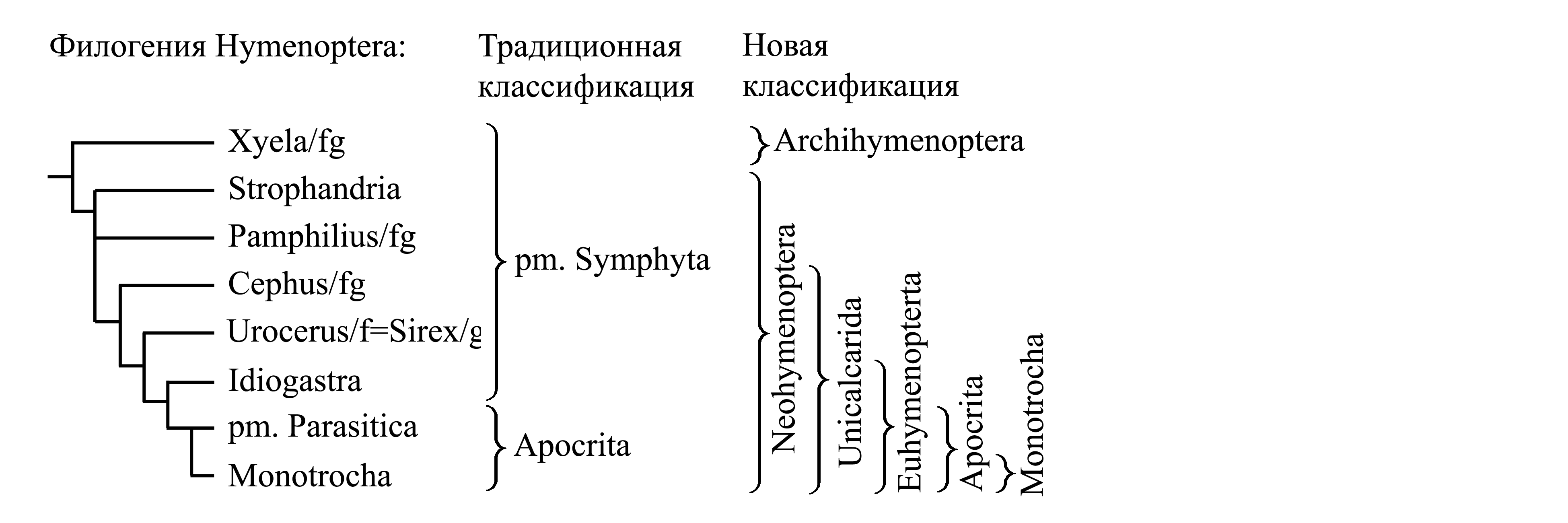
1. Плезиоморфон Symphyta Gerstaecker 1867 NomCirc. – сидячебрюхие {synn.circ: Ventrosessiles Latreille 1802b = Sessiliventres Latreille 1804c = Serricaudia Dumeril 1805 = Uroprista Dumeril 1805 = Phytiphaga Latreille 1807 = Pristuria Rafinesque 1815 = Securifera Latreille 1817 = Phytophaga Billberg 1820 (non Phytophaga Dumeril 1805) = Serrifera Berthold 1827 = Phytospheces Leunis 1860 = Chalastogastra Konow 1897 = Prohymenoptera Crampton 1916a}. В отличие от стебельчатобрюхих, первый сегмент брюшка не слит с заднегрудью и не отделен от следующего сегмента перетяжкой (рис. 10.7.1). Личинки питаются растениями – либо живыми частями растений (пилильщики и др.), либо древесиной, разрушаемой грибами (рогохвосты и др.). Личинки сохраняют способность перемещаться в поисках пищи – либо имеют развитые ноги и ложноножки (открытоживущие личинки пилильщиков – рис. 10.7.3), либо безногие, обитающие в тканях растений. Наиболее важные таксоны сидячебрюхих перепончатокрылых:
1.1. Xyelidae, или Archihymenoptera Grimaldi & Engel 2005 NomCirc. {nom.typ.: Xyela/fg [f:1834; g:1819]} (рис. 10.7.1). Мелкие, менее 5 мм. Яйцеклад длинный, выступает далеко за вершину брюшка. Личинки развиваются либо в мужских стробилах сосен, либо в почках и молодых побегах хвойных, либо на листьях вязовых и ореховых. Archihymenoptera являются наиболее древними среди перепончатокрылых, известны начиная с триаса. Включают около 50 современных видов, обитают только в Голарктике (преимущественно в Неарктике). По мнению разных авторов, Archihymenoptera – это либо голофилетический таксон сестринский всем остальным перепончатокрылым (которых в этом случае объединяют в таксон Neohymenoptera Grimaldi & Engel 2005), либо парафилетический таксон, предковый для прочих перепончатокрылых.
|
1.2. Tenthredinoidea, или Strophandria Ross 1937 NomCirc. – пилильщики {nom.typ.: Tenthredo/fg [f:1802; g:L.1758(gen.214)] (incl. Cimbex, Diprion, Arge)}. Название «пилильщики» связано с тем, что яйцеклад пиловидно зазубрен и обычно используется для того, чтобы сделать надрез в листе и вставить в него яйцо (которое таким образом оказывается закрепленным и обеспеченным влагой для своего развития). Личинки, называемые ложногусеницами, похожи на гусениц бабочек: удлиненные, сегменты груди и брюшка сходного размера и строения; ноги развиты (см. Metabola (1Cc); второй и последующие сегменты брюшка несут по паре ложноножек, так что бывают развиты 8 или меньшее число пар ложноножек; ложноножки расположены на одной линии с грудными ногами (рис. 10.7.3A). Личинки многих видов живут открыто на растениях, питаясь их зелеными частями; у некоторых видов личинки постоянно или временно обитают внутри растений или в галлах. Включает около 7000 видов, т.е. большую часть видов сидячебрюхих; распространены всесветно. WWWWWWWWWWWWWWWWWWWWWWW |
|
1.3. Pamphiliidae – паутинные пилильщики {nom.typ.: Pamphilius/fg [f:1890; g:1802]}. Личинки сходны с ложногусеницами пилильщиков. Около 250 видов в Голарктике.
1.4. Cephidae – стеблевые пилильщики {nom.typ.:Cephus/fg [f:1834; g:1802]}. Личинки безногие, развиваются внутри стеблей растений. Около 100 видов.
1.5. Uroceridae (или "Siricidae") – рогохвостовые (nom.typ.: Urocerus/f=Sirex/g [f:1802; g:1761] (incl. Xiphydria)}. Личинки живут в древесине и питаются благодаря симбиозу с грибом, которым самка заражает дерево. В брюшке взрослой самки имеется пара мешковидных органов – микангиев (mycangia), открывающихся протоками в основании яйцеклада. У личинки самки в складке между первым и вторым сегментами брюшка имеется пара углублений, выстланных железистым эпителием и служащих для культивирования гриба. При развитии имаго из личинки споры культивируемого гриба попадают в микангии. При откладке яиц самка выделяет придаточными железами яйцеклада (гомологичными ядовитым железам жалящих) секрет, который вместе со спорами гриба инъецируется в древесину. Разные виды рогохвостовых используют разные виды грибов, относящихся с аскомицетам и базидиомицетам (Gauld & Bolton 1988). Около 200 видов.
1.6. Orussidae, или Idiogastra Rohwer & Cushman 1917 NomCirc. {syn.circ.: Protoparasitica Engel 2005; nom.typ.: Orussus/fg [f:1834; g:1796]}. У самки яйцеклад очень длинный, в покое свернут внутри брюшка; приспособлен для откладки яиц в древесину. Личинки безногие, развиваются в ходах, проложенных в мертвой древесине другими насекомыми; могут питаться как эктопаразитоиды на личинках жуков и рогохвостов; они также могут питаться разлагающейся древесиной, подобно личинкам рогохвостов (Gauld & Bolton 1988). Около 75 видов, преимущественно в южном полушарии (Vilhelmsen 2003).
Оруссиды представляют собой переходную форму между рогохвостами (относящимися к плезиоморфону Symphyta) и наездниками (относящимися к Apocrita). В соответствии с этим разные авторы либо относят оруссид к рогохвостами, либо объединяют оруссид с Apocrita в таксон Euhymenoptera Engel & Grimaldi 2004 NomCirc..
2. Apocrita Gerstaecker 1867 NomCirc. {syn.circ.: Petiolata Sharp 1894; nom.typ.: Vespa/f=Cynips/g (sine Orussus). Голофилетический таксон, характеризуется следующими аутапоморфиями.
У имаго и куколки тергит первого сегмента брюшка слит с грудью (стернит первого сегмента не выражен, как и у прочих Hymenoptera); основание второго сегмента брюшка суженное и очень подвижно сочленено с первым сегментом (рис. 10.7.4D). Таким образом кажущаяся граница между «грудью» и «брюшком» проходит по узкому подвижному сочленению между первым и вторым сегментами брюшка. Отдел тела, кажущийся «грудью» и образованный тремя грудными сегментами и истинным первым сегментом брюшка, называется мезосома (mesosoma); истинный первый сегмент брюшка, слитый с грудью и кажущийся ее частью, называется промежуточный сегмент, или проподеум (propodeum); сочленение между ним и истинным вторым сегментом брюшка называется стебелек (petiolus); брюшко начиная с истинного второго сегмента называется метаcома (metasoma). Стебелек исходно возник как приспособление к паразитоидному развитию личинок (см. ниже): наличие стебелька позволяет самке манипулировать брюшком при нападении на подвижное насекомое, парализовать его уколом яйцеклада и отложить на него яйца. При этом яд, используемый для парализации, выделяется придаточной железой. У жалящих перепончатокрылых (Monotrocha) стебелек способствует использованию жала, которое представляет собой видоизмененный яйцеклад и служит для парализации или умерщвления добычи, которой самка выкармливает личинок, а также для защиты потомства. Стебельчатое строение брюшка свойственно обоим полам, хотя используется только самкой. Стебельчатое брюшко сохраняется у всех Apocrita, в том числе и у тех, которые его не используют и утратили яйцеклад.
Личинки всех Apocrita совершенно безногие. Исходно личинка Apocrita ведет паразитоидный образ жизни: питается на живом насекомом-хозяине (подобно паразиту), а в конце своего развития убивает и доедает его (подобно хищнику). Жертвами (или, как их часто называют, хозяевами) в большинстве случаев являются личинки и куколки насекомых с полным превращением, неподвижные или малоподвижные имаго некоторых Plantisuga и яйца различных насекомых и пауков. Паразитоидное развитие свойственно большинству видов Apocrita. В наиболее примитивном случае самка при откладке яйца парализует хозяина уколом яйцеклада, а личика развивается на поверхности хозяина как эктопаразитоид. При этом иногда жертва остается парализованной на протяжении всего питания личинки. Иногда парализация временная, в таком случае личинка питается на жертве, ведущей активный образ жизни. Иногда парализации вообще не происходит, так что самка откладывает яйцо на активное насекомое. В некоторых таксонах Apocrita личинки адаптировались к жизни внутри тела насекомого (эндопаразитоиды). У разных видов паразитоидов, в зависимости от соотношения размеров паразитоида и жертвы, на одной жертве может развиваться одна или много личинок. Личинки некоторых Apocrita перешли от паразитоидного развития на насекомых-галлообразователях к самостоятельному галлообразованию, то есть являются фитофагами. Некоторые представители Apocrita перешли от паразитоидного образа жизни к выкармливанию личинок в гнезде (см. ниже, Monotrocha).
У большинства личинок Apocrita полости средней кишки (энтодермальной) и задней кишки (эктодермальной) разобщены до конца периода питания, так что выделение экскрементов происходит только перед линькой на куколку; это связано с тем, что личинки Apocrita исходно питаются жидкой гемолимфой насекомых. Лишь у некоторых пчелиных кишечник стал вторично сквозным начиная с первого личиночного возраста (все пчелиные перешли к питанию пыльцой). В отличие от Apocrita, у Idiogastra сохраняется связь средней и задней кишки, несмотря на паразитоидный образ жизни.
|
2.1. Плезиоморфон Parasitica Hartig 1837 NomCirc. {synn. circ.: Pedonculiventres Latreille 1804c = Spiculifera Westwood 1840 = Entomospheces Leunis 1860 = Tristega Konov 1897; nom.typ.: Ichneumon/f=Cynips/g [f:1802; g:1758(gen.213)]}. Личинки большинства видов являются паразитоидами на насекомых (см. выше); личинки некоторых видов паразитируют на кладках яиц насекомых, т.е. формально являются хищниками. В некоторых таксонах Parasitica личинки перешли от паразитоидного питания на насекомых-галлообразователях к самостоятельному образованию галлов на растениях и питанию тканями галла, т.е. являются настоящими фитофагами. Включает порядка 65 000 описанных видов, т.е. бóльшую часть всех видов Hymenoptera. WWWWWWWWWWWWWWWWWWWW |
 |
2.2. Monotrocha Hartig 1837 NomCirc. – жалящие {по циркумскрипции соответствует: «Aculeata» auct. (non Scopoli 1763); nom.typ.: Vespa/f=Sphex/g [f:1781; g:1758(gen.216)]}. Характеризуется следующими апоморфиями. У самки имаго VIII и IX сегменты брюшка сильно модифицированы, с вестигиальными тергитами, втянуты в VII сегмент брюшка (рис. 10.7.4D); находящийся на них яйцеклад превращен в выдвигающееся жало и утратил функцию проведения яиц. Вертлужок утрачен. Антенны у самки исходно 12-члениковые, у самца исходно 13-члениковые (в некоторых таксонах на 1–2 членика меньше). Включает несколько десятков тысяч видов.
Исходным для Monotrocha, как и для Parasitica, является паразитоидный образ жизни личинок и временная парализация жертвы при откладке яиц (см. выше). Многие представители Monotrocha перешли от паразитоидного образа жизни, исходно свойственного Apocrita (см. выше) к выкармливанию личинок в специально построенном гнезде. В этом случае самка парализует жертву не временно, а постоянно, и не только откладывает яйцо на жертву, но и перемещает эту жертву из ее естественного места обитания в гнездо, приспособленное для развития личинки. В наиболее примитивном случае самка лишь перемещает парализованную добычу в естественное укрытие и откладывает на нее яйцо, так что далее личинка развивается как эктопаразитоид. Во многих случаях самка сначала делает гнездо определенной видоспецифичной конструкции, затем разыскивает добычу, парализует ее, перемещает в гнездо, откладывает на жертву яйцо и запечатывает гнездо. У некоторых видов самка помещает в гнездо не одно, а несколько парализованных насекомых; таким образом, здесь, в отличие от исходного паразитоидного питания, возможно питание личинок более мелкими жертвами. У некоторых видов самка не запечатывает гнездо, а на протяжении всего развития личинок выкармливает их свежеубитыми насекомыми или иной пищей животного происхождения. В отдельных случаях произошел переход от выкармливания личинок животной пищей к выкармливанию иной пищей: в частности пчелиные (Apidae, или Apoidea) и цветочные осы – (Masaridae) выкармливают личинок не насекомыми, а пыльцой растений; большинство муравьев (Formicidae) выкармливают личинок насекомыми, но некоторые муравьи перешли к выкармливанию личинок семенами растений, а муравьи-листорезы (Attini) кормят личинок специально разводимыми грибами. Во всех случаях все функции, связанные заботой о потомстве (постройка гнезда, добывание пищи, защита незапечатанного гнезда и др.) выполняются только самкой; несмотря на то, что в некоторых случаях эти работы бывают трудоемкими, самцы никогда не принимают в них участия; то есть настоящих семей у перепончатокрылых не бывает (в отличие от некоторых других насекомых со сложным поведением, у которых возникают постоянные семейные пары). Вероятно, это вызвано тем, что исходно основные родительские функции связаны с наличием жалящего яйцеклада, который используется и как средство добывания пищи для личинок, и как средство их защиты, а яйцеклад может быть развит только у самок.
Вместо этого среди Monotrocha, выкармливающих личинок в гнезде, имеется большое число социальных видов. Одна и та же особая форма социальности, присущая Monotrocha, встречается у представителей разных таксонов, то есть возникала в ходе эволюции независимо несколько раз. При этой форме социальности все самки колонии разделены на плодовитую касту (царицы) и бесплодную касту (рабочие); в отличие от других социальных животных (термиты и др.), у социальных перепончатокрылых на касты делятся только самки, тогда как все самцы являются обычными плодовитыми особями и не участвуют в функционировании колонии. Многократное возникновение такой социальности вызвано сочетанием двух причин: 1) для выполнения сложных функций по выращиванию потомства (добыванию пищи, защиты и др.) самкам выгодно кооперироваться друг с другом, поскольку самцы в этом участвовать не могут; 2) как и все перепончатокрылые, эти насекомые имеют гапло-диплоидное определение пола, и поэтому самка имеет бóльшее генетическое сходство со своими сестрами, чем с матерью (см. выше). Это приводит к тому, что естественный отбор может быть направлен на поддержание такого поведения, которое способствует выращиванию своих сестер в ущерб выращиванию своих дочерей.
* * *
KlugeIMGP0616-Vesp.JPG)
Подробнее см. Н.Ю. Клюге 2020. «Систематика насекомых и принципы кладоэндезиса».
