CLADOENDESIS OF EPHEMEROPTERA
|
CLADOENDESIS OF EPHEMEROPTERA |
||||
| zzz | ||||
|
|
|
Tetramerotarsata, or Baetis/fg2
(Panephemeroptera![]() Euephemeroptera
Euephemeroptera![]() Euplectoptera
Euplectoptera![]() Anteritorna
Anteritorna![]() pm.Tridentiseta
pm.Tridentiseta![]()
- Tetramerotarsata)
Nomen hierarchicum: ![]() Baetis/fg2
[fg:1815] (sine. Siphlonurus, Dipteromimus, Ameletus,
Metretopus, Acanthametropus, Ametropus, Metamonius, Oniscigaster,
Ameletopsis, Rallidens; incl. Siphlaenigma)
Baetis/fg2
[fg:1815] (sine. Siphlonurus, Dipteromimus, Ameletus,
Metretopus, Acanthametropus, Ametropus, Metamonius, Oniscigaster,
Ameletopsis, Rallidens; incl. Siphlaenigma)
Nomen circumscribens:
![]()
![]()
![]() Tetramerotarsata
Kluge 1997c: 528.
Tetramerotarsata
Kluge 1997c: 528.
In circumscription fits:
— fam. Baetidae: McCafferty & Edmunds 1979: 6
— superfam. Baetoidea: Kluge & Studemann & Landolt & Gonser 1995: 107
— superfam. Tetramerotarsata Kluge 1997c: 528
— Tetramerotarsata = Baetis/fg2: Kluge 2000: 248
References. Kluge 1997c:
![]() *
* ![]() *;
– Staniczek 1997:
*;
– Staniczek 1997:
![]() *
* ![]() *
*
![]() *;
– Kluge 2004:
*;
– Kluge 2004: ![]() *
*
![]() *
* ![]() * – Kluge
& Novikova 2011:
* – Kluge
& Novikova 2011: ![]() *.
*.
|
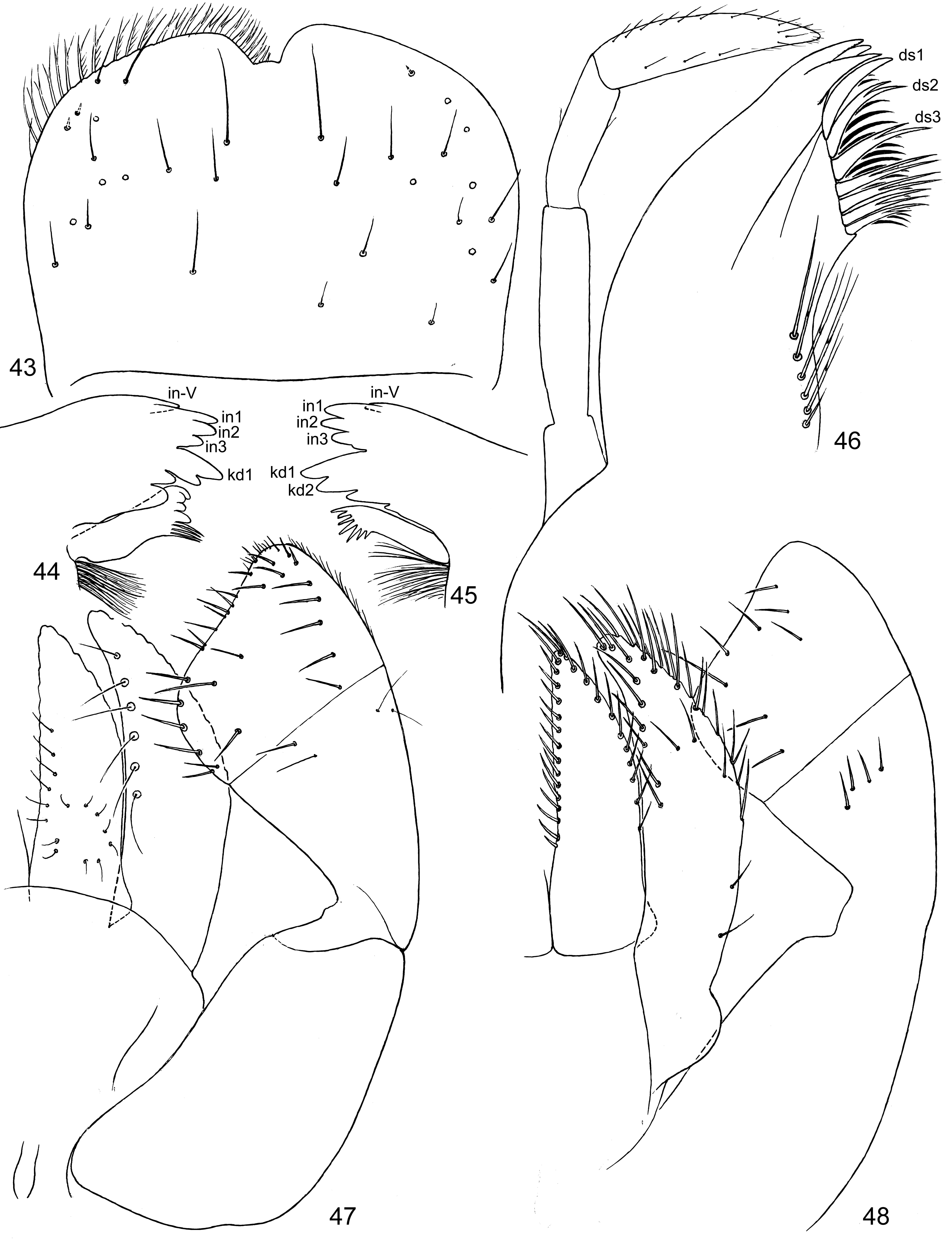
Autapomorphies of Tetramerotarsata. (1) Imaginal and subimaginal tarsus of middle and hind legs consists of 4 segments only, including 1st segment fused with tibia; thus, only 3 segments are movable (Kluge 2004: Fig.27:E) (unlike initial for mayflies 5-segmented tarsus with 4 segments movable). Most probably, tarsi became 4-segmented because their initial 1st and 2nd tarsomeres are completely fused together, without any traces of boundary between them. This can be revealed from the following fact: certain tarsomeres are ventrally-apically produced into a thorn; number of such thorn-bearing tarsomeres is species-specific; in some species of Liberevenata fore tarsus of female, which has 5 tarsomeres, has the same number of thorn-bearing tarsomeres as middle and hind tarsi (which always have 4 tarsomeres); in all these cases position of the thorn-bearing tarsomeres indicates that the 1st tarsomere of 4-segmented middle and hind tarsus corresponds to the 1st and 2nd tarsomeres of 5-segmented fore tarsus. In order to indicate serial homology between tarsomeres of different pairs of legs, tarsomeres of the 4-segmented middle and hind tarsus should be termed not "1st–4th", but 1st+2nd, 3rd, 4th and 5th. This apomorphy is nearly unique among Ephemeroptera. In some other Ephemeroptera reduction of tarsal segments takes place in connection with general degeneration of legs (in Prosopistoma/f1=g2, Geminovenata, some Fossoriae); unlike them, in Tetramerotarsata legs are always normally developed, functional, with well-developed long tarsi. Among all mayflies, only in Machadorythus tarsi have the same structure as in Tetramerotarsata (see Index of characters [2.2.83]). In other respects legs of Tetramerotarsata are non-modified or variously modified in subordinate taxa. On middle and hind legs the 1st (immobile) tarsal segment is not shortened, often the longest. Fore tarsus of male retains all 5 segments, the 1st segment being either non-shortened (in Siphlaenigma), or strongly shortened [see Liberevenata (3) below]. Fore tarsus of female is either generally the same as in male – 5 segments are retained, 1st of which is shortened; or (e.g. janae [Siphlaenigma] and costai [Indobaetis]) fore tarsus of female is 4-segmented, like middle and hind tarsi. Fore tarsus of male never has apical thorns. On other legs, maximum number of apical thorns is 3; in this case, on 5-segmented fore leg of female 2nd, 3rd and 4th tarsomeres are thorn-bearing (Kluge & Novikova 2011: Fig. 4), and on 4-segmentede middle and hind legs of both sexes 1st+2nd, 3rd and 4th tarsomeres are thorn-bearing (Kluge & Novikova 2011: Fig. 5). In certain taxa number of apical thorns is less than 3; on fore leg of female it can be either the same as on middle and hind legs, or different (see Index of characters [2.2.78]). Claws of all legs of imago and subimago of both sexes are always ephemeropteroid. (2) Imaginal and subimaginal mesonotum has sublateroscutal suture – a paired transverse suture, which crosses sublateroscutum and separates dorsal bases of anterior and posterior scuto-coxal muscles. At the same time, portion of sublateroscutum behind the sublateroscutal suture is not separated from the posterior scutal protuberance (which bears base of scuto-lateropostnotal muscle) and together with it looks as an integral piece (Kluge 2004: Fig.26:A, 29:A). Sublateroscutal suture is a new formation, not occurring in other Ephemeroptera (it should not be confused with the transverse interscutal suture of Caenoptera and some Tricorythus/fg1, which passes not anteriad, but posteriad of the base of the posterior scuto-coxal muscle). (3) Imaginal and subimaginal anterior paracoxal suture is transferred anteriorly, thus anepisternum is smaller than katepisternum (Kluge 2004: Fig.26:A, 27:F, 29:A). Non-unique apomorphy (see Index of characters [2.2.19]). Unlike Rallidens, Heptagennota and Furcatergaliae, anterior paracoxal suture is complete. (4) Mesonotal suture is lost because its left and right halves are curved backward, elongated and fused medially with the median longitudinal suture and laterally with lateroparapsidal suture. The conclusion that mesonotal suture had disappeared by this way, is based on the fact that subimago of Siphlaenigma retains a vestige of mesonotal suture: it has a form of a pair of longitudinal lines, which go close to the median longitudinal suture and separate a narrow light bold area lying mediad of these lines, from pigmented microtrichia-bearing area lying laterad of them (Kluge 2004: Fig.26:A). In all other Tetramerotarsata traces of mesonotal suture are lost and subimaginal mesonotum is entirely covered with microtrichia. Non-unique apomorphy; independently from Tetramerotarsata mesonotal suture disappeared by the same way in Leptophlebia/fg1 and some Fimbriatotergaliae (see Index of characters [2.2.8]). (5) On fore wing CuP is basally curved and strongly diverges from CuA. This curvation is retained in primitive representatives of Tetramerotarsata – Siphlaenigma and Palaeocloeon (Kluge 2004: Fig.27:D), while in Turbanoculata the transverse basal portion of CuP is lost, so that base of CuP stands apart from base of CuA. Non-unique apomorphy; the same in Leptophlebia/fg1 only. (6) Labrum with following features: Outer side with a transverse row of branching setae close to distal margin [in Siphlaenigma this row is regular in its lateral parts and irregular in median part, in Turbanoculata it is entirely regular – see Turbanoculata (4) below]; at a distance from distal margin a pair of long submedian setae are present. In cases of strong specialization of labrum, these details can be lost. (7) On maxilla apical-ventral row of setae is transferred to ventral side; it is well-developed in Siphlaenigma (Kluge 2004: Fig.26:B), being vestigial or lost in Turbanoculata. (8) Paraglossa with a longitudinal row of setae on dorsal side and initially with a longitudinal row of setae on ventral side (Kluge 2004: Fig.26:D, 28:C). The dorsal row is always retained, but the ventral row is secondarily lost in many Turbanoculata. (?) Labial palp initially has 2nd segment widened apically, ... Characters of Tetramerotarsata of unclear phylogenetic status. (9) On fore wing MP2 has a form of intercalary, i.e. lost connection with MP1 (Kluge 2004: Fig.27:A) (but at the same time begins at a distance from wing base, unlike Posteritorna and Caenoptera). Non-unique apomorphy, in many other taxa disjunction of MP2 occurs as individual variability. (10) Cubital field of fore wing [see Anteritorna (1)] initially with 2 veins arising from CuA, the proximal (posterior) of them being bifurcate; this condition can be retained Siphlaenigma (Penniket 1962: Fig.1); in Liberevenata these 2 veins become simple intercalaries; posterior of them can be short, in Palaeocloeon seems to be lost (Kluge 2004: Fig.27:A). Non-unique apomorphy (see Index of characters [2.2.52]). (11) Larval claw on inner side has 2 longitudinal rows of denticles. Such claw structure is initial for Tetramerotarsata, being present in Siphlaenigma and many Turbanoculata. Non-unique apomorphy (see Index of characters [1.2.21]). In some Tetramerotarsata denticles of that row, which is situated nearer to posterior side of the claw, are diminished or lost, thus only one row is present, being situated nearer to anterior side of the claw (anterior is the side directed anteriorly when the leg is directed by apex laterally and by knee dorsally); similar single anterior-inner row of denticles occurs also in some of that mayfly taxa, which initially have one row of denticles – Pentamerotarsata, Ephemerella/fg1, Leptophlebia/fg1 (see Index of characters [1.2.21]). In selected species of Tetramerotarsata (belonging both to taxa with two rows and taxa with one row of denticles) the denticles can be secondarily lost. (12) Tergalius has anal rib situated on anal margin (unlike Siphlonurus/fg1 and some others, which have anal rib at a distance from anal margin – see Index of characters [1.3.28]). Costal margin of tergalius is armed by costal rib, thus tergalius often has an integral rib entirely bordering its margins. In a few taxa (particularly, in Cloeon/fg1 and Callibaetis) anal rib is completely lost. In one species – itajara [Cloeodes] – each tergalius, besides costal and anal ribs, has additional poorly developed middle rib in front of main trachea. (13) Larval antennae are elongate, usually much longer than head; only in selected specialized species antennae are not longer than head (while in majority of other mayflies antennae are short, subequal or slightly longer than head). Plesiomorphies of Tetramerotarsata. Larval head always retains hypognathous position (see below, Variable characters of Turbanoculata). Maxilla [see (7)] normally has 3 canines and 3 dentisetae, which are different from more proximal setae of the same (inner-dorsal) row [see Anteritorna (2) and Plesiomorphies of Tridentiseta] (Kluge 2004: Fig.26:B, 28:B,D Kluge 12: Fig.27; Kluge 2012: Fig.5; Kluge 2015: Fig.5; Kluge 2016: Fig.2; Kluge & Novikova 2016: Fig.36, 70, 123; Kluge & Novikova 2017: Fig.77; Kluge 2017: Fig.8, 109; Kluge & al. 2017: Fig6); in selected individuals seta proximad of 3rd dentiseta has the same shape as 2nd an 3rd dentisetae, so total number of dentisetae looks as increased to 4 (Kluge & al. 2017: Fig7); sometimes other setae of this (inner-dorsal) row have similar shape (Kluge 2016: Fig. 46); in peculiar case number of dentisetae is diminished to 2 (see Index of characters [1.1.37]). |
Size. Usually small, fore wing length 2–12 mm.
Age and distribution. Late Cretaceous (see Palaeocloeon) – Recent; world-wide.
| The taxon Tetramerotarsata (or Baetis/fg2) is divided into: |
1. Siphlaenigma
![]()
2. Liberevenata,
or Baetis/fg3 ![]()
2.1. plesiomorphon
Palaeocloeon ![]()
2.2.
Turbanoculata, or Baetis/fg4 ![]()
{kind=link}